

Akt-dependent regulation of NF-{kappa}B is controlled by mTOR and Raptor in association with IKK
- PMID: 18519641
- PMCID: PMC2418585
- DOI: 10.1101/gad.1662308
Akt-dependent regulation of NF-{kappa}B is controlled by mTOR and Raptor in association with IKK
Abstract
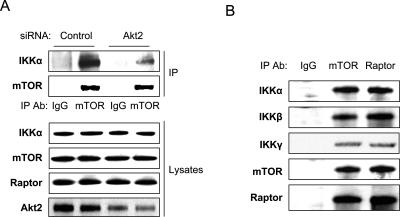
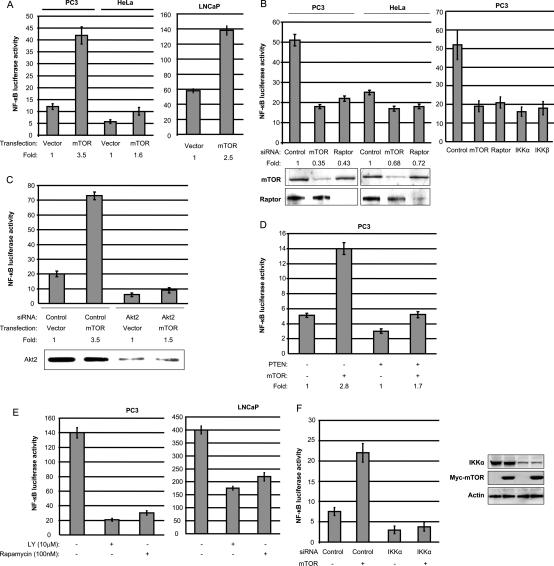
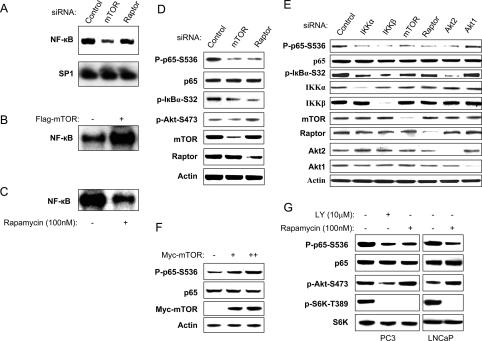
While NF-kappaB is considered to play key roles in the development and progression of many cancers, the mechanisms whereby this transcription factor is activated in cancer are poorly understood. A key oncoprotein in a variety of cancers is the serine-threonine kinase Akt, which can be activated by mutations in PI3K, by loss of expression/activity of PTEN, or through signaling induced by growth factors and their receptors. A key effector of Akt-induced signaling is the regulatory protein mTOR (mammalian target of rapamycin). We show here that mTOR downstream from Akt controls NF-kappaB activity in PTEN-null/inactive prostate cancer cells via interaction with and stimulation of IKK. The mTOR-associated protein Raptor is required for the ability of Akt to induce NF-kappaB activity. Correspondingly, the mTOR inhibitor rapamycin is shown to suppress IKK activity in PTEN-deficient prostate cancer cells through a mechanism that may involve dissociation of Raptor from mTOR. The results provide insight into the effects of Akt/mTOR-dependent signaling on gene expression and into the therapeutic action of rapamycin.
Figures




References
-
- Agarwal A., Das K., Lerner N., Sathe S., Cicek M., Casey G., Sizemore N. The Akt/IKK pathway promotes angiogenic/metastatic gene expression in colorectal cancer by activating NF-κB and β-catenin. Oncogene. 2005;24:1021–1031. - PubMed
-
- Basseres D., Baldwin A. NF-κB and IKK pathways in oncogenic initiation and progression. Oncogene. 2006;25:6817–6830. - PubMed
-
- Bhaskar P.T., Hay N. The two TORCs and Akt. Dev. Cell. 2007;12:487–502. - PubMed
-
- Bonizzi G., Karin M. The two NF-κB activation pathways and their roles in innate and adaptive immunity. Trends Immunol. 2004;25:280–288. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous