

Desmin mediates TNF-alpha-induced aggregate formation and intercalated disk reorganization in heart failure
- PMID: 18519735
- PMCID: PMC2396798
- DOI: 10.1083/jcb.200710049
Desmin mediates TNF-alpha-induced aggregate formation and intercalated disk reorganization in heart failure
Abstract
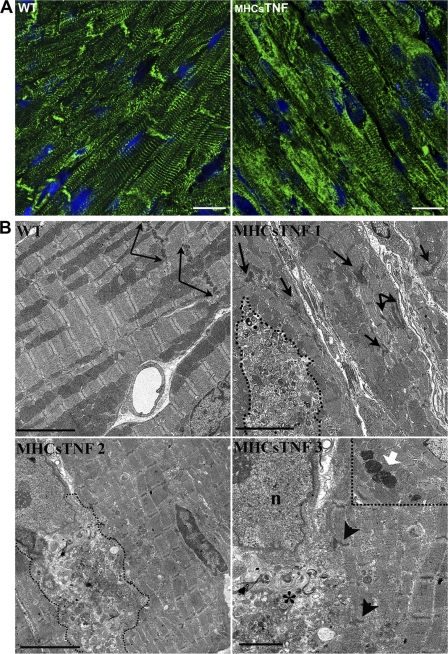
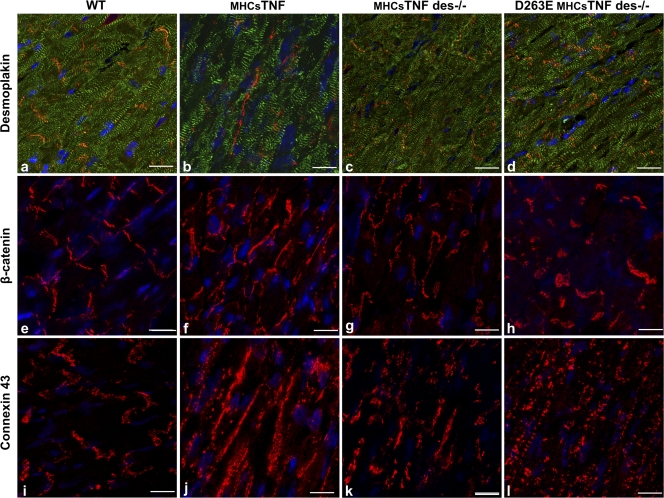
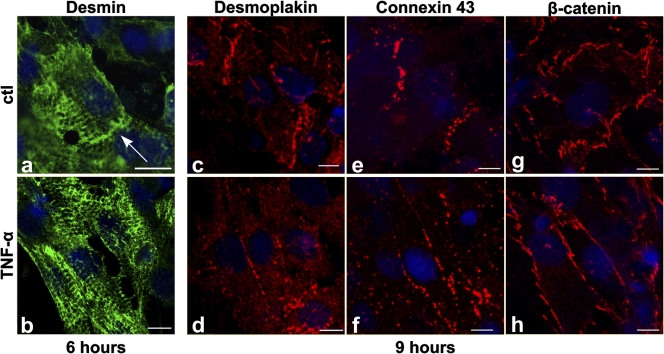
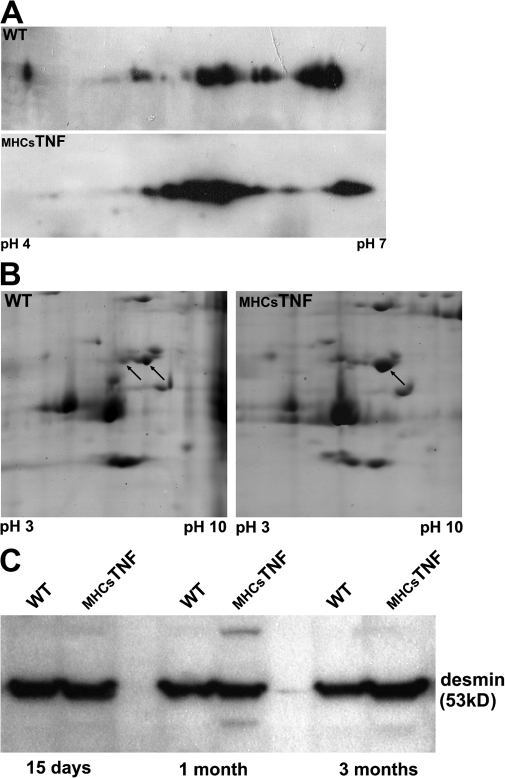
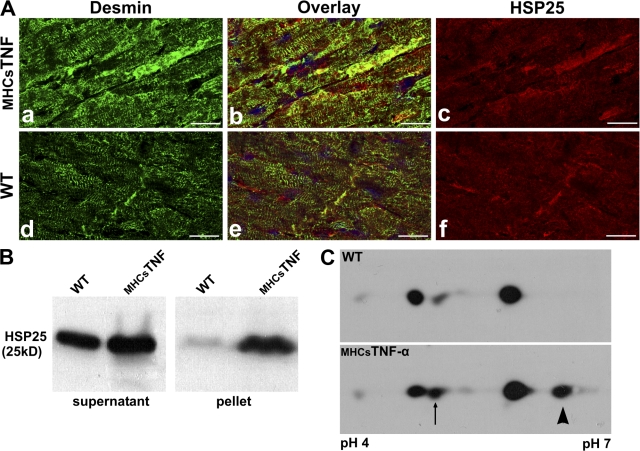
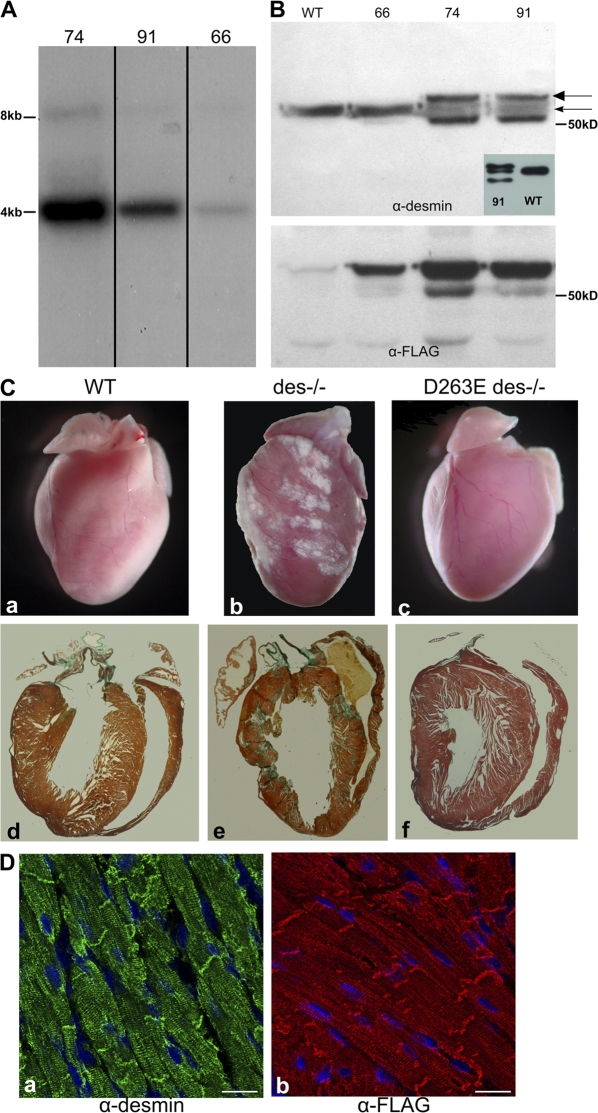
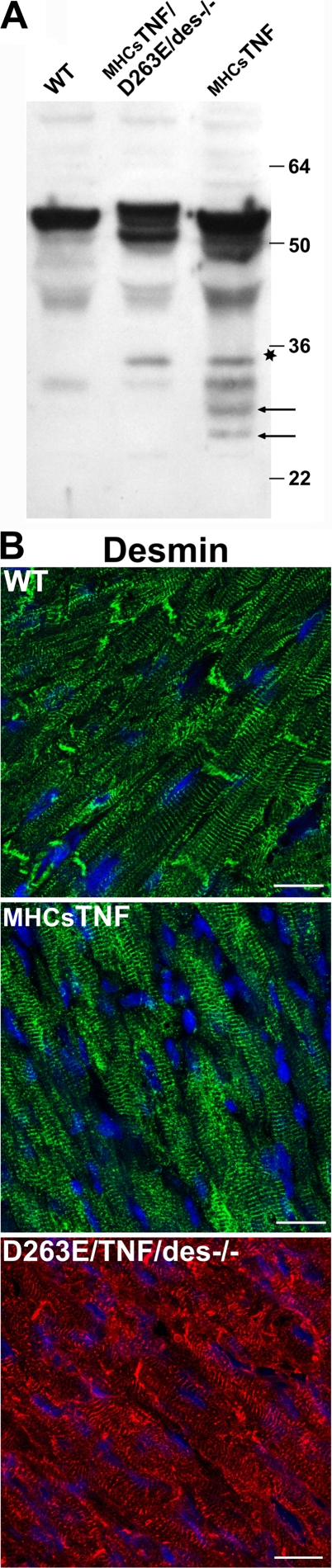
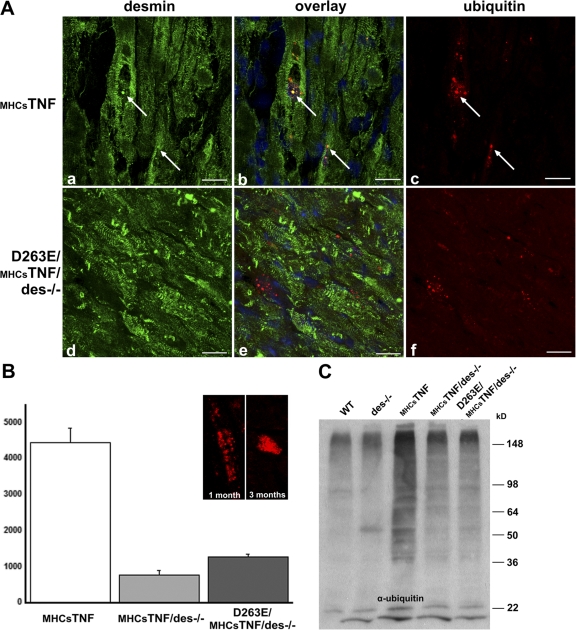
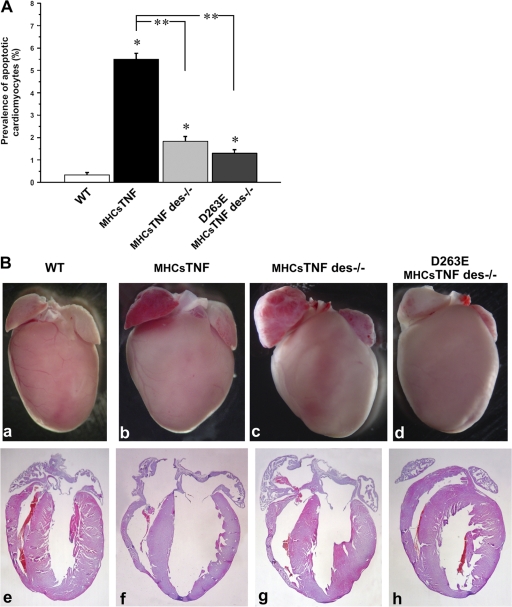
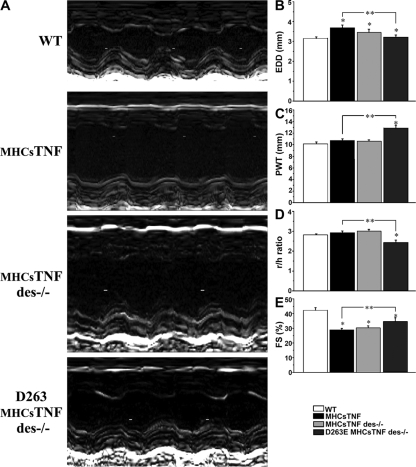
We explored the involvement of the muscle-specific intermediate filament protein desmin in the model of tumor necrosis factor alpha (TNF-alpha)-induced cardiomyopathy. We demonstrate that in mice overexpressing TNF-alpha in the heart (alpha-myosin heavy chain promoter-driven secretable TNF-alpha [MHCsTNF]), desmin is modified, loses its intercalated disk (ID) localization, and forms aggregates that colocalize with heat shock protein 25 and ubiquitin. Additionally, other ID proteins such as desmoplakin and beta-catenin show similar localization changes in a desmin-dependent fashion. To address underlying mechanisms, we examined whether desmin is a substrate for caspase-6 in vivo as well as the implications of desmin cleavage in MHCsTNF mice. We generated transgenic mice with cardiac-restricted expression of a desmin mutant (D263E) and proved that it is resistant to caspase cleavage in the MHCsTNF myocardium. The aggregates are diminished in these mice, and D263E desmin, desmoplakin, and beta-catenin largely retain their proper ID localization. Importantly, D263E desmin expression attenuated cardiomyocyte apoptosis, prevented left ventricular wall thinning, and improved the function of MHCsTNF hearts.
Figures
References
-
- Bajaj, G., and R.K. Sharma. 2006. TNF-alpha-mediated cardiomyocyte apoptosis involves caspase-12 and calpain. Biochem. Biophys. Res. Commun. 345:1558–1564. - PubMed
-
- Bar, H., S.V. Strelkov, G. Sjoberg, U. Aebi, and H. Herrmann. 2004. The biology of desmin filaments: how do mutations affect their structure, assembly, and organisation? J. Struct. Biol. 148:137–152. - PubMed
-
- Bence, N.F., R.M. Sampat, and R.R. Kopito. 2001. Impairment of the ubiquitin-proteasome system by protein aggregation. Science. 292:1552–1555. - PubMed
-
- Bennett, E.J., N.F. Bence, R. Jayakumar, and R.R. Kopito. 2005. Global impairment of the ubiquitin-proteasome system by nuclear or cytoplasmic protein aggregates precedes inclusion body formation. Mol. Cell. 17:351–365. - PubMed
-
- Brancolini, C., A. Sgorbissa, and C. Schneider. 1998. Proteolytic processing of the adherens junctions components beta-catenin and gamma-catenin/plakoglobin during apoptosis. Cell Death Differ. 5:1042–1050. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
