

Roles of Oxa1-related inner-membrane translocases in assembly of respiratory chain complexes
- PMID: 18522806
- PMCID: PMC2658530
- DOI: 10.1016/j.bbamcr.2008.05.004
Roles of Oxa1-related inner-membrane translocases in assembly of respiratory chain complexes
Abstract
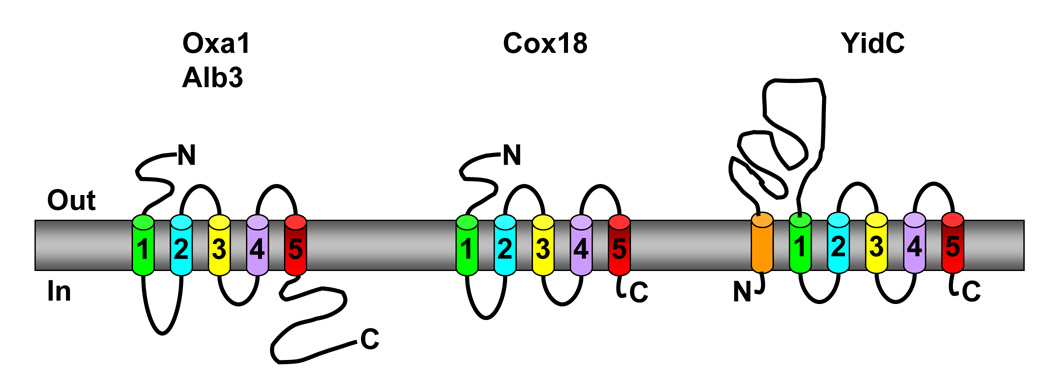
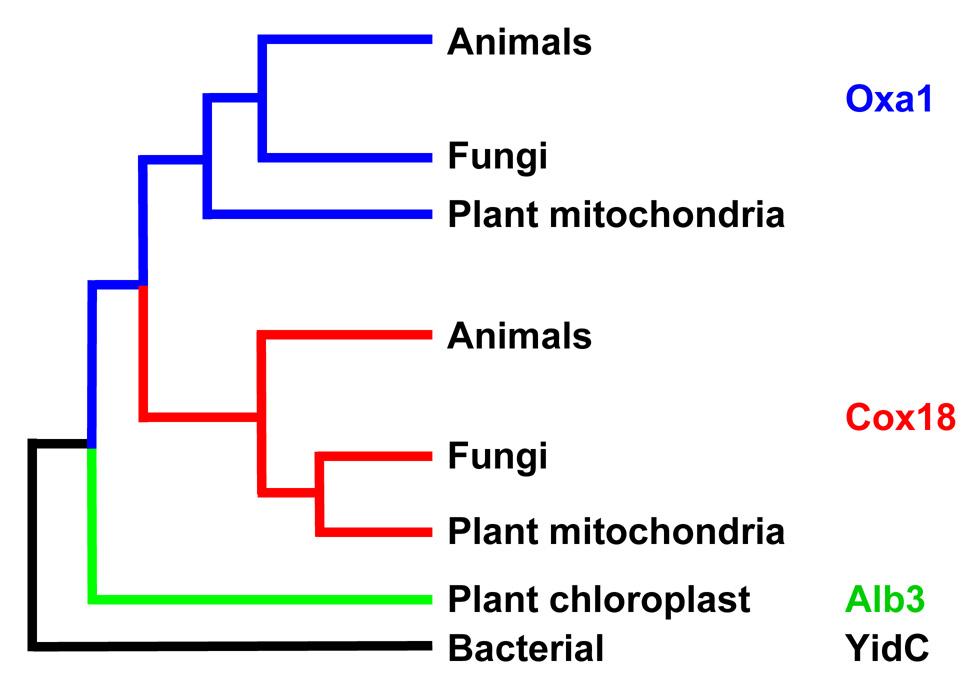
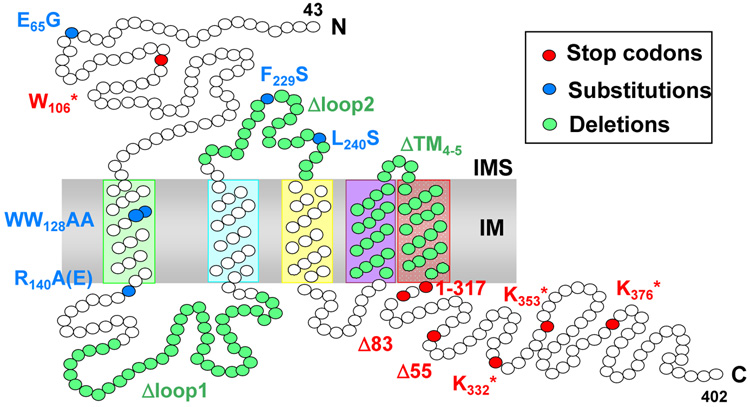
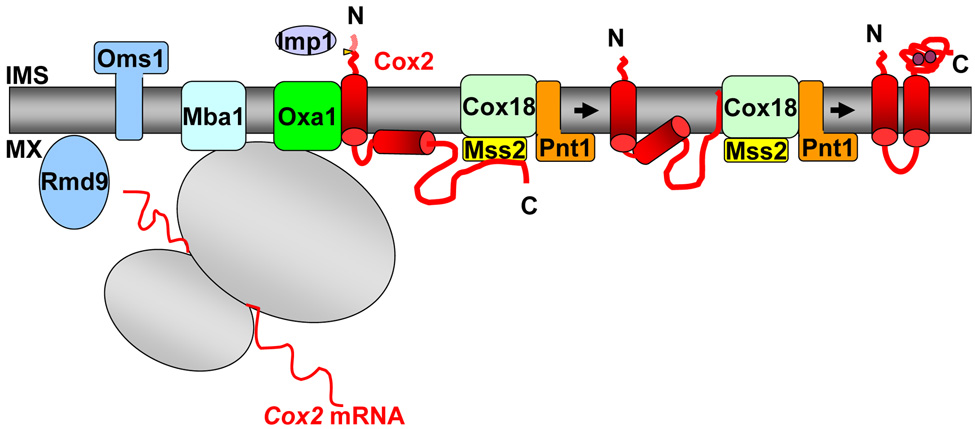
Members of the family of the polytopic inner membrane proteins are related to Saccharomyces cerevisiae Oxa1 function in the assembly of energy transducing complexes of mitochondria and chloroplasts. Here we focus on the two mitochondrial members of this family, Oxa1 and Cox18, reviewing studies on their biogenesis as well as their functions, reflected in the phenotypic consequences of their absence in various organisms. In yeast, cytochrome c oxidase subunit II (Cox2) is a key substrate of these proteins. Oxa1 is required for co-translational translocation and insertion of Cox2, while Cox18 is necessary for the export of its C-terminal domain. Genetic and biochemical strategies have been used to investigate the functions of distinct domains of Oxa1 and to identify its partners in protein insertion/translocation. Recent work on the related bacterial protein YidC strongly indicates that it is capable of functioning alone as a translocase for hydrophilic domains and an insertase for TM domains. Thus, the Oxa1 and Cox18 probably catalyze these reactions directly in a co- and/or posttranslational way. In various species, Oxa1 appears to assist in the assembly of different substrate proteins, although it is still unclear how Oxa1 recognizes its substrates, and whether additional factors participate in this beyond its direct interaction with mitochondrial ribosomes, demonstrated in S. cerevisiae. Oxa1 is capable of assisting posttranslational insertion and translocation in isolated mitochondria, and Cox18 may posttranslationally translocate its only known substrate, the Cox2 C-terminal domain, in vivo. Detailed understanding of the mechanisms of action of these two proteins must await the resolution of their structure in the membrane and the development of a true in vitro mitochondrial translation system.
Figures
References
-
- Dolezal P, Likic V, Tachezy J, Lithgow T. Evolution of the molecular machines for protein import into mitochondria. Science. 2006;313:314–318. - PubMed
-
- Herrmann JM, Neupert W. Protein insertion into the inner membrane of mitochondria. IUBMB Life. 2003;55:219–225. - PubMed
-
- Lang BF, Burger G, O'Kelly CJ, Cedergren R, Golding GB, Lemieux C, Sankoff D, Turmel M, Gray MW. An ancestral mitochondrial DNA resembling a eubacterial genome in miniature. Nature. 1997;387:493–497. - PubMed
-
- Yi L, Dalbey RE. Oxa1/Alb3/YidC system for insertion of membrane proteins in mitochondria, chloroplasts and bacteria. Mol. Membr. Biol. 2005;22:101–111. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
