

Neonatal chimerization with human glial progenitor cells can both remyelinate and rescue the otherwise lethally hypomyelinated shiverer mouse
- PMID: 18522848
- PMCID: PMC3358921
- DOI: 10.1016/j.stem.2008.03.020
Neonatal chimerization with human glial progenitor cells can both remyelinate and rescue the otherwise lethally hypomyelinated shiverer mouse
Abstract
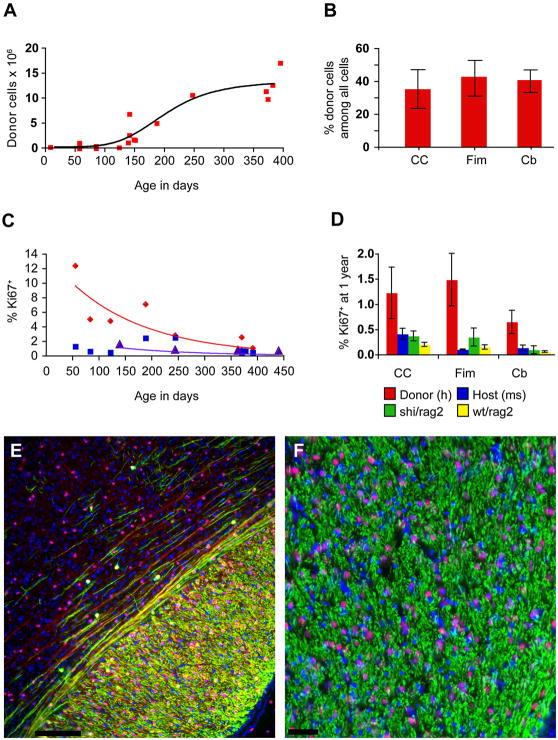
Congenitally hypomyelinated shiverer mice fail to generate compact myelin and die by 18-21 weeks of age. Using multifocal anterior and posterior fossa delivery of sorted fetal human glial progenitor cells into neonatal shiverer x rag2(-/-) mice, we achieved whole neuraxis myelination of the engrafted hosts, which in a significant fraction of cases rescued this otherwise lethal phenotype. The transplanted mice exhibited greatly prolonged survival with progressive resolution of their neurological deficits. Substantial myelination in multiple regions was accompanied by the acquisition of normal nodes of Ranvier and transcallosal conduction velocities, ultrastructurally normal and complete myelination of most axons, and a restoration of a substantially normal neurological phenotype. Notably, the resultant mice were cerebral chimeras, with murine gray matter but a predominantly human white matter glial composition. These data demonstrate that the neonatal transplantation of human glial progenitor cells can effectively treat disorders of congenital and perinatal hypomyelination.
Figures
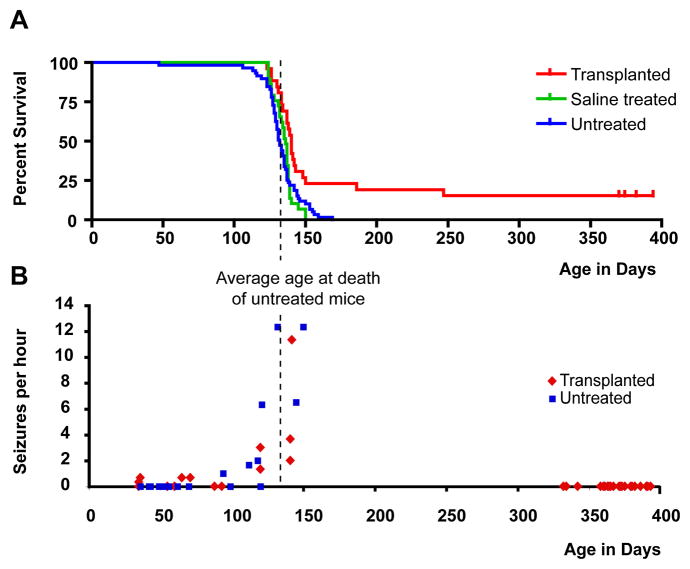

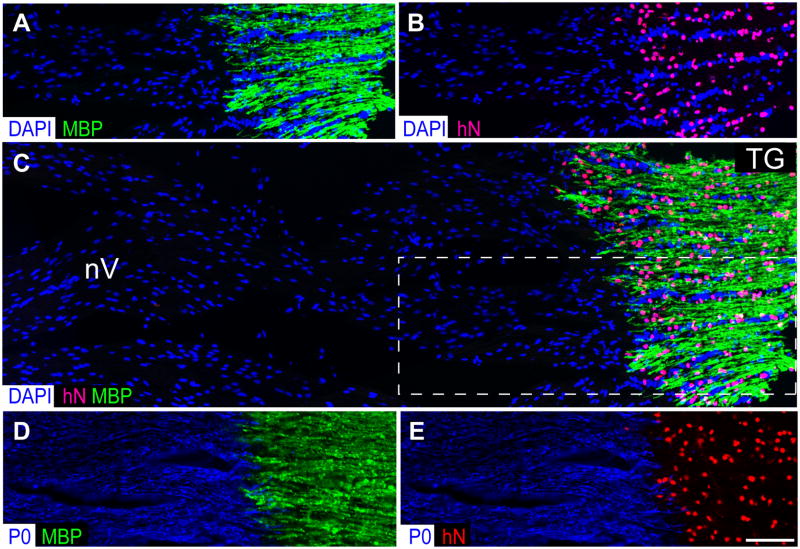
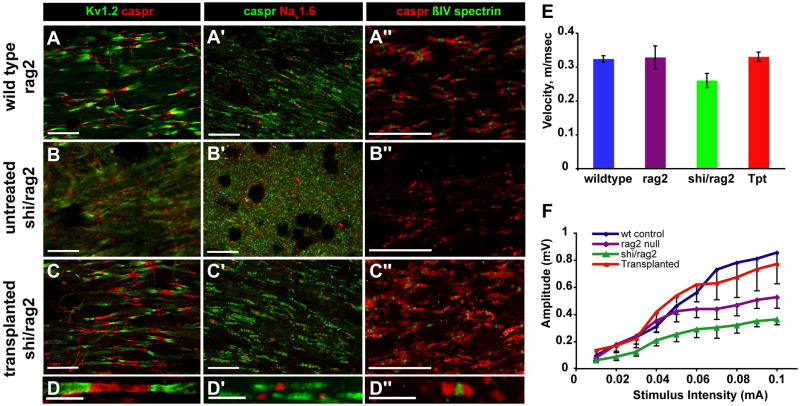
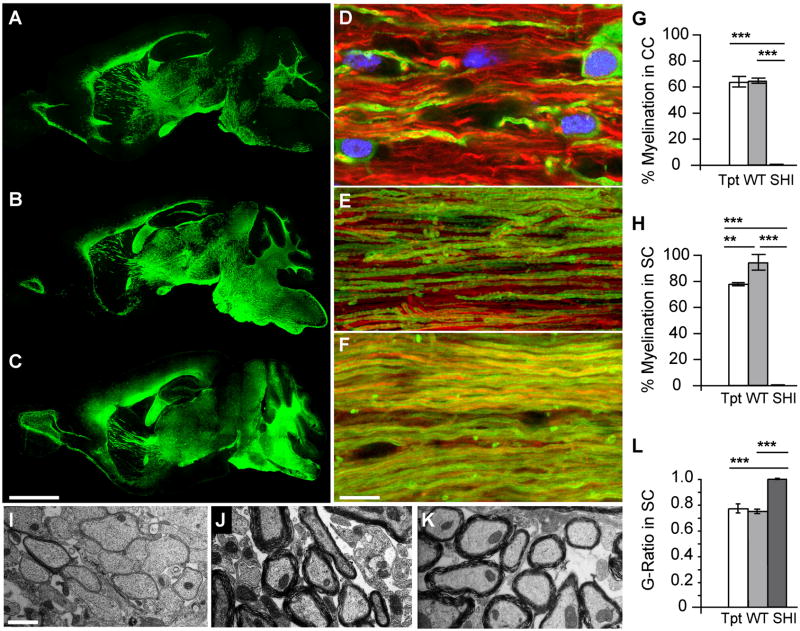
Comment in
-
Rescue of congenital hypomyelination by progenitor cell transplantation.Cell Stem Cell. 2008 Jun 5;2(6):519-20. doi: 10.1016/j.stem.2008.05.017. Cell Stem Cell. 2008. PMID: 18522843
References
-
- Archer D, Cuddon P, Lipsitz D, Duncan I. Myelination of the canine central nervous sytem by glial cell transplantation: A model for repair of human myelin disease. Nature Medicine. 1997;3:54–59. - PubMed
-
- Archer DR, Leven S, Duncan ID. Myelination by cryopreserved xenografts and allografts in the myelin-deficient rat. Exp Neurol. 1994;125:268–277. - PubMed
-
- Deguchi K, Oguchi K, Takashima S. Characteristic neuropathology of leukomalacia in extremely low birth weight infants. Pediatr Neurol. 1997;16:296–300. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
