

The p110gamma isoform of phosphatidylinositol 3-kinase regulates migration of effector CD4 T lymphocytes into peripheral inflammatory sites
- PMID: 18523230
- PMCID: PMC2516906
- DOI: 10.1189/jlb.0807561
The p110gamma isoform of phosphatidylinositol 3-kinase regulates migration of effector CD4 T lymphocytes into peripheral inflammatory sites
Abstract
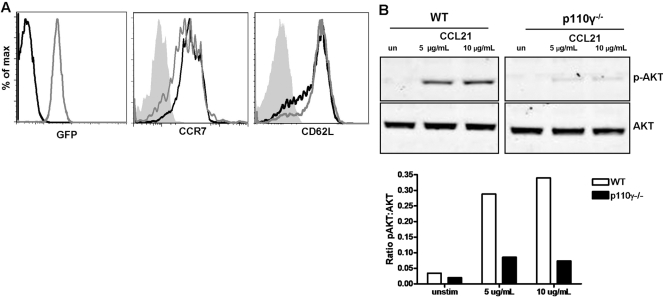
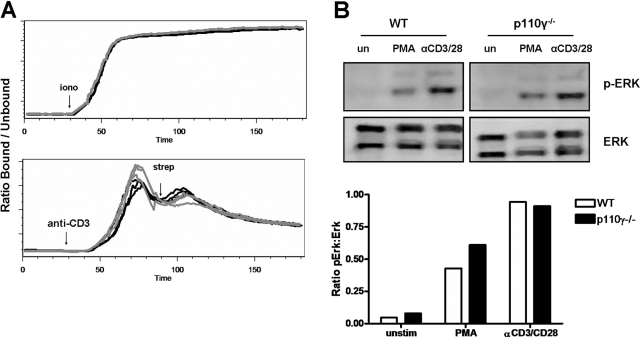
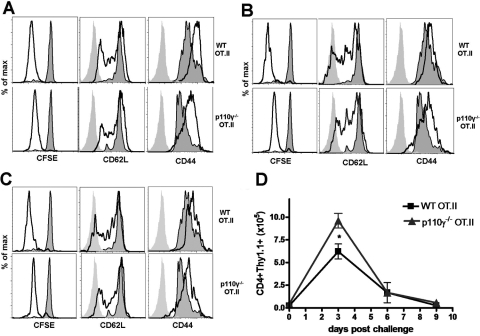
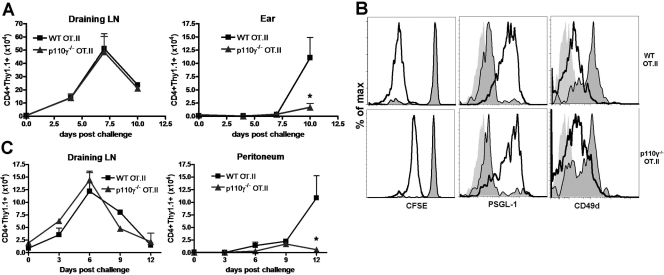
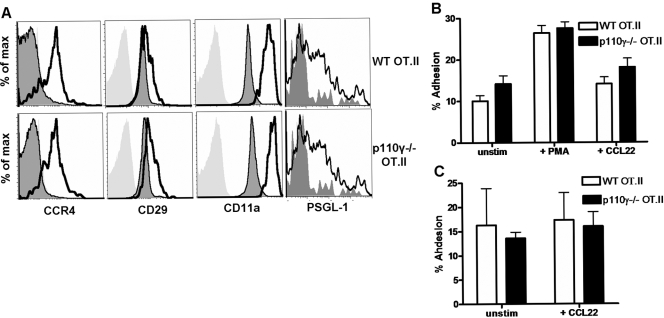
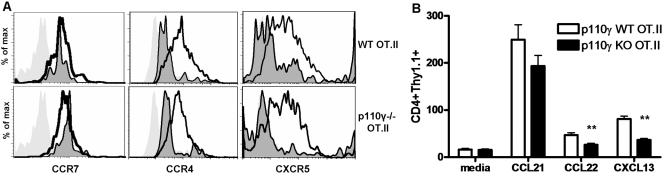
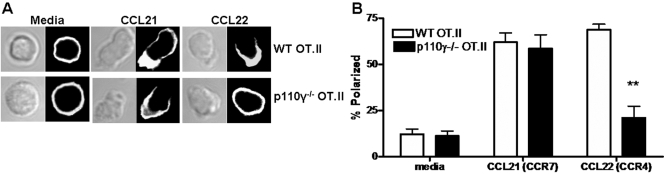
The role of PI-3K in leukocyte function has been studied extensively. However, the specific role of the p110gamma isoform of PI- 3K in CD4 T lymphocyte function has yet to be defined explicitly. In this study, we report that although p110gamma does not regulate antigen-dependent CD4 T cell activation and proliferation, it plays a crucial role in regulating CD4 effector T cell migration. Naïve p110gamma(-/-) CD4 lymphocytes are phenotypically identical to their wild-type (WT) counterparts and do not exhibit any defects in TCR-mediated calcium mobilization or Erk activation. In addition, p110gamma-deficient CD4 OT.II T cells become activated and proliferate comparably with WT cells in response to antigen in vivo. Interestingly, however, antigen-experienced, p110gamma-deficient CD4 OT.II lymphocytes exhibit dramatic defects in their ability to traffic to peripheral inflammatory sites in vivo. Although antigen-activated, p110gamma-deficient CD4 T cells express P-selectin ligand, beta2 integrin, beta1 integrin, CCR4, CXCR5, and CCR7 comparably with WT cells, they exhibit impaired F-actin polarization and migration in response to stimulation ex vivo with the CCR4 ligand CCL22. These findings suggest that p110gamma regulates the migration of antigen-experienced effector CD4 T lymphocytes into inflammatory sites during adaptive immune responses in vivo.
Figures
References
-
- Ward S G. T lymphocytes on the move: chemokines, PI 3-kinase and beyond. Trends Immunol. 2006;27:80–87. - PubMed
-
- Rommel C, Camps M, Ji H. PI3K δ and PI3K γ: partners in crime in inflammation in rheumatoid arthritis and beyond? Nat Rev Immunol. 2007;7:191–201. - PubMed
-
- Wymann M P, Bjorklof K, Calvez R, Finan P, Thomast M, Trifilieff A, Barbier M, Altruda F, Hirsch E, Laffargue M. Phosphoinositide 3-kinase γ: a key modulator in inflammation and allergy. Biochem Soc Trans. 2003;31:275–280. - PubMed
-
- Fukao T, Terauchi Y, Kadowaki T, Koyasu S. Role of phosphoinositide 3-kinase signaling in mast cells: new insights from knockout mouse studies. J Mol Med. 2003;81:524–535. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
