

The central memory CD4+ T cell population generated during Leishmania major infection requires IL-12 to produce IFN-gamma
- PMID: 18523296
- PMCID: PMC2614376
- DOI: 10.4049/jimmunol.180.12.8299
The central memory CD4+ T cell population generated during Leishmania major infection requires IL-12 to produce IFN-gamma
Abstract
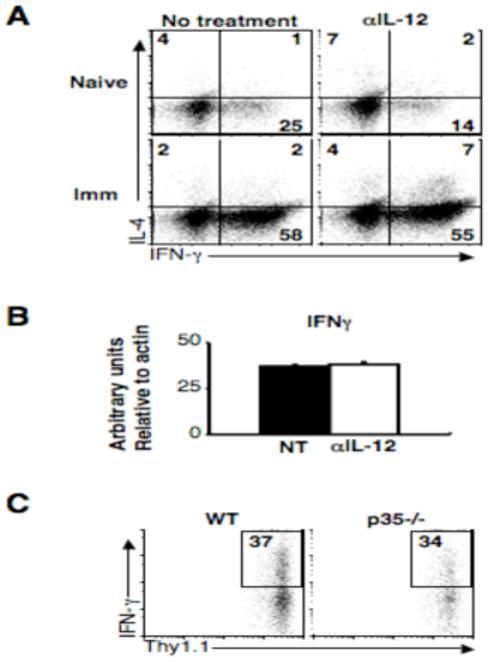
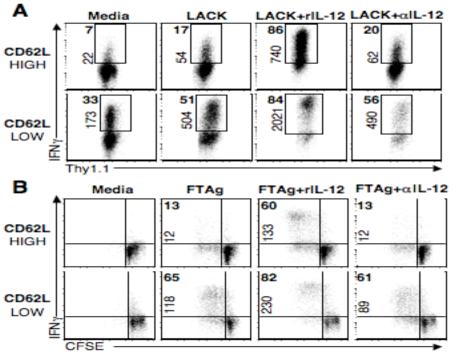
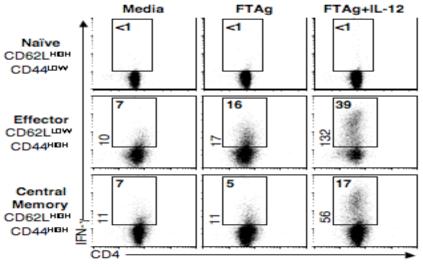
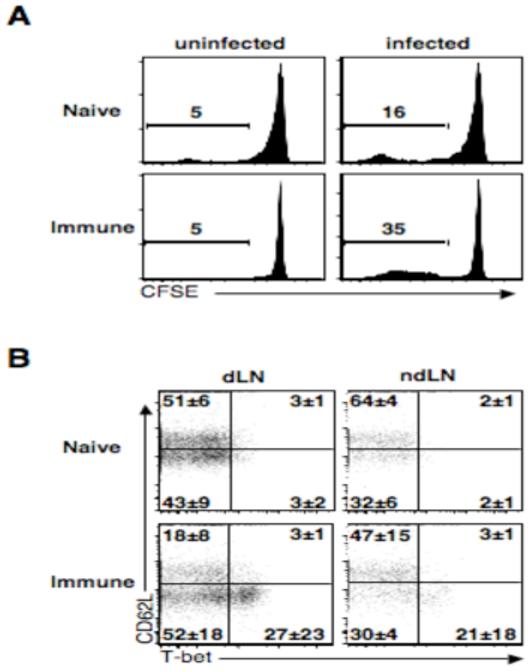
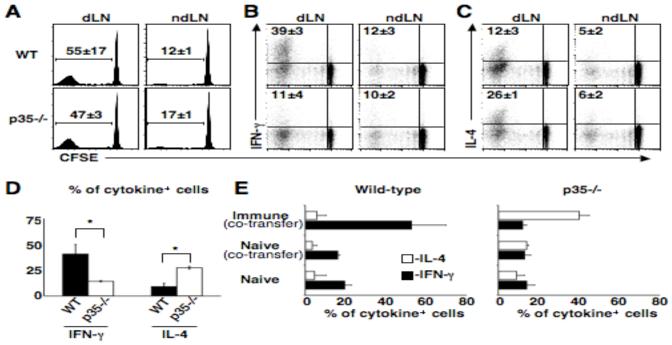
Central memory CD4(+) T cells provide a pool of lymph node-homing, Ag-experienced cells that are capable of responding rapidly after a secondary infection. We have previously described a population of central memory CD4(+) T cells in Leishmania major-infected mice that were capable of mediating immunity to a secondary infection. In this study, we show that the Leishmania-specific central memory CD4(+) T cells require IL-12 to produce IFN-gamma, demonstrating that this population needs additional signals to develop into Th1 cells. In contrast, effector cells isolated from immune mice produced IFN-gamma in vitro or in vivo in the absence of IL-12. In addition, we found that when central memory CD4(+) T cells were adoptively transferred into IL-12-deficient hosts, many of the cells became IL-4 producers. These studies indicate that the central memory CD4(+) T cell population generated during L. major infection is capable of developing into either Th1 or Th2 effectors. Thus, continued IL-12 production may be required to ensure the development of Th1 cells from this central memory T cell pool, a finding that has direct relevance to the design of vaccines dependent upon central memory CD4(+) T cells.
Figures
References
-
- Scott P, Artis D, Uzonna J, Zaph C. The development of effector and memory T cells in cutaneous leishmaniasis: the implications for vaccine development. Immunol Rev. 2004;201:318–338. - PubMed
-
- Sacks D, Noben-Trauth N. The immunology of susceptibility and resistance to Leishmania major in mice. Nat Rev Immunol. 2002;2:845–858. - PubMed
-
- Sallusto F, Lenig D, Forster R, Lipp M, Lanzavecchia A. Two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nature. 1999;401:708–712. - PubMed
-
- Zaph C, Uzonna J, Beverley SM, Scott P. Central memory T cells mediate long-term immunity to Leishmania major in the absence of persistent parasites. Nat Med. 2004;10:1104–1110. - PubMed
-
- Park AY, Hondowicz B, Kopf M, Scott P. The role of IL-12 in maintaining resistance to Leishmania major. J Immunol. 2002;168:5771–5777. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
