

Epidural stimulation induced modulation of spinal locomotor networks in adult spinal rats
- PMID: 18524907
- PMCID: PMC2904311
- DOI: 10.1523/JNEUROSCI.0080-08.2008
Epidural stimulation induced modulation of spinal locomotor networks in adult spinal rats
Abstract
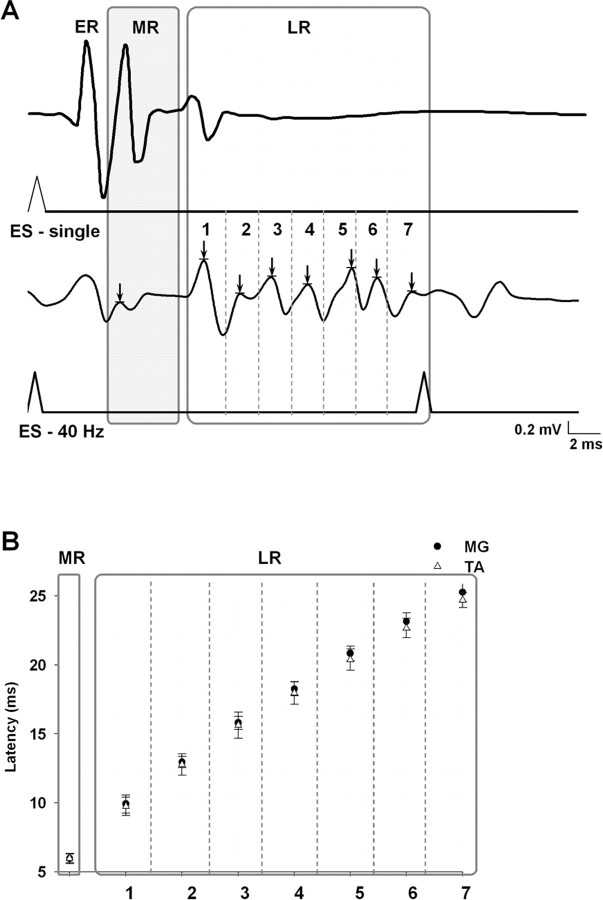
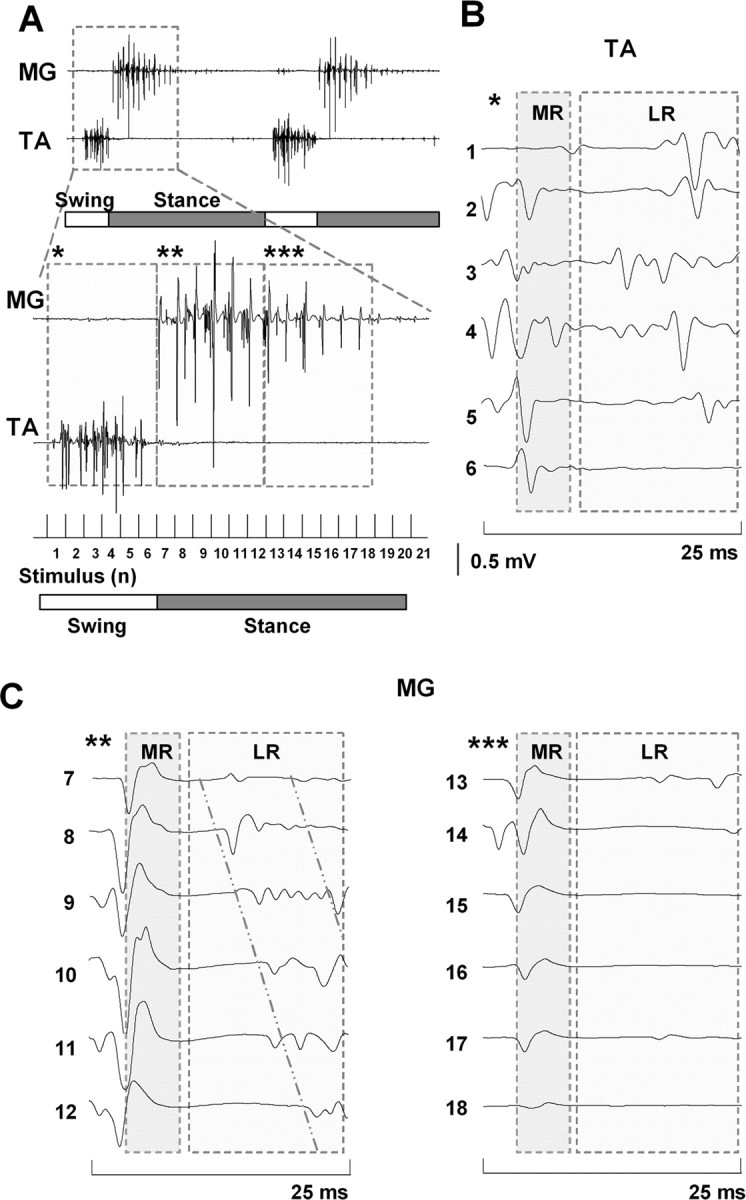
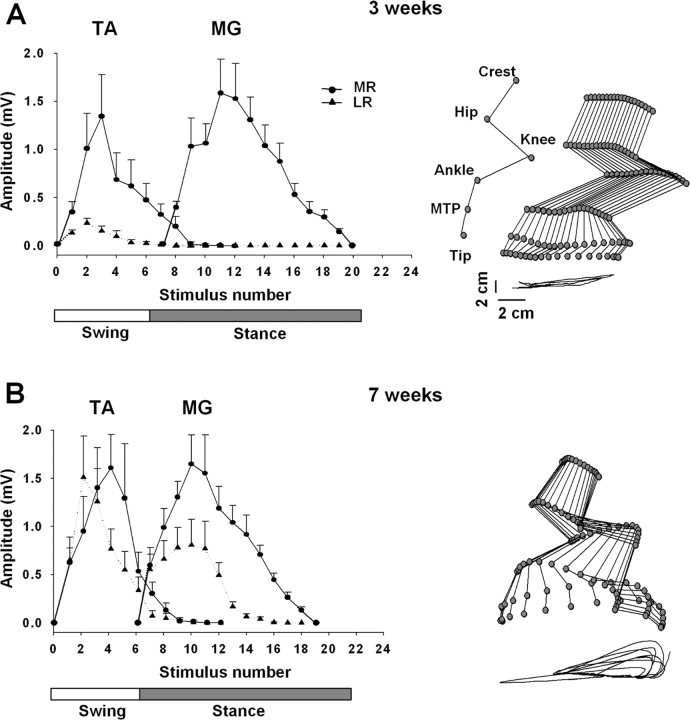
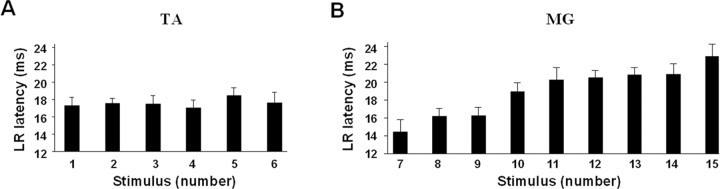
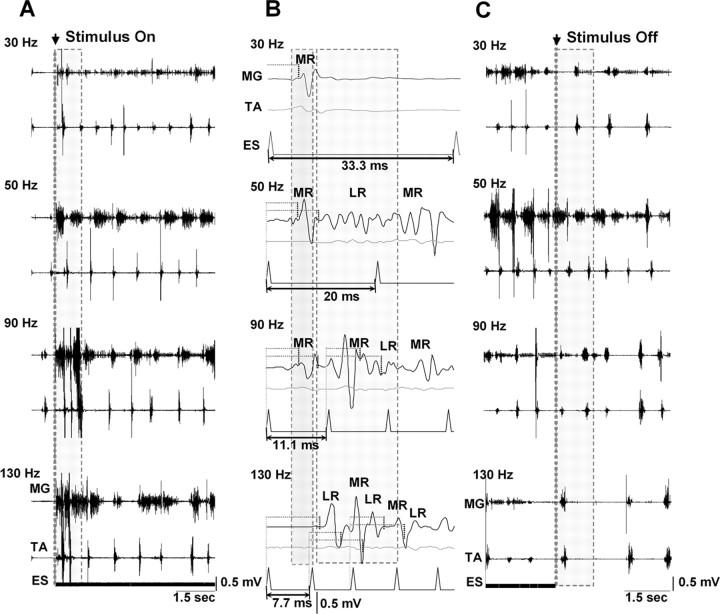
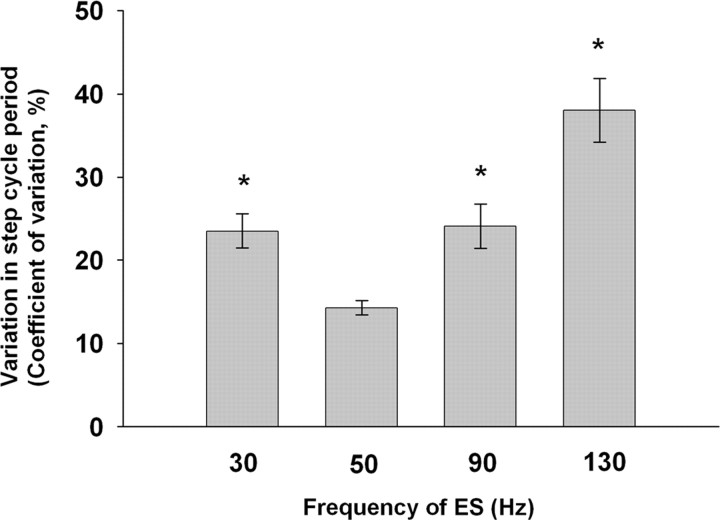
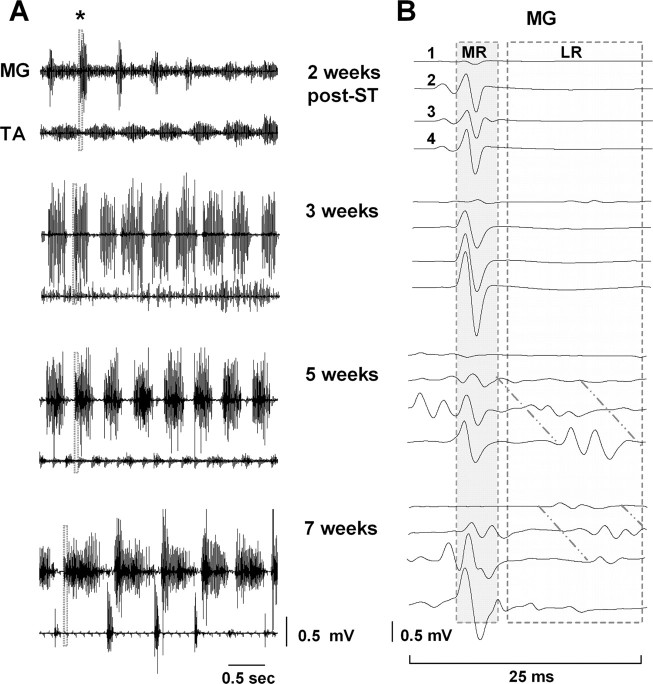
The importance of the in vivo dynamic nature of the circuitries within the spinal cord that generate locomotion is becoming increasingly evident. We examined the characteristics of hindlimb EMG activity evoked in response to epidural stimulation at the S1 spinal cord segment in complete midthoracic spinal cord-transected rats at different stages of postlesion recovery. A progressive and phase-dependent modulation of monosynaptic (middle) and long-latency (late) stimulation-evoked EMG responses was observed throughout the step cycle. During the first 3 weeks after injury, the amplitude of the middle response was potentiated during the EMG bursts, whereas after 4 weeks, both the middle and late responses were phase-dependently modulated. The middle- and late-response magnitudes were closely linked to the amplitude and duration of the EMG bursts during locomotion facilitated by epidural stimulation. The optimum stimulation frequency that maintained consistent activity of the long-latency responses ranged from 40 to 60 Hz, whereas the short-latency responses were consistent from 5 to 130 Hz. These data demonstrate that both middle and late evoked potentials within a motor pool are strictly gated during in vivo bipedal stepping as a function of the general excitability of the motor pool and, thus, as a function of the phase of the step cycle. These data demonstrate that spinal cord epidural stimulation can facilitate locomotion in a time-dependent manner after lesion. The long-latency responses to epidural stimulation are correlated with the recovery of weight-bearing bipedal locomotion and may reflect activation of interneuronal central pattern-generating circuits.
Figures
References
-
- Barbeau H, Rossignol S. Initiation and modulation of the locomotor pattern in the adult chronic spinal cat by noradrenergic, serotonergic and dopaminergic drugs. Brain Res. 1991;546:250–260. - PubMed
-
- Barthélemy D, Leblond H, Rossignol S. Characteristics and mechanisms of locomotion induced by intraspinal microstimulation and dorsal root stimulation in spinal cats. J Neurophysiol. 2007;97:1986–2000. - PubMed
-
- Burke RE, Degtyarenko AM, Simon ES. Patterns of locomotor drive to motoneurons and last-order interneurons: clues to the structure of the CPG. J Neurophysiol. 2001;86:447–462. - PubMed
-
- Chau C, Barbeau H, Rossignol S. Effects of intrathecal alpha1- and alpha2-noradrenergic agonists and norepinephrine on locomotion in chronic spinal cats. J Neurophysiol. 1998;79:2941–2963. - PubMed
-
- Chiba A, Oshio K, Inase M. Magnetically evoked EMGs in rats. Neurol Res. 2003;25:87–91. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical