

Selective ligand recognition by a diversity-generating retroelement variable protein
- PMID: 18532877
- PMCID: PMC2408619
- DOI: 10.1371/journal.pbio.0060131
Selective ligand recognition by a diversity-generating retroelement variable protein
Abstract
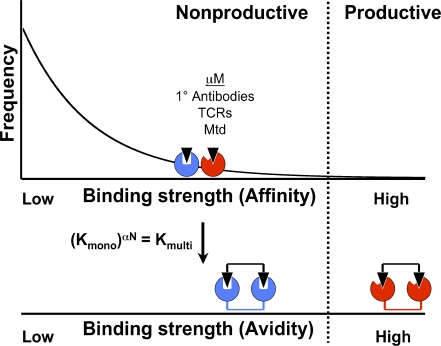
Diversity-generating retroelements (DGRs) recognize novel ligands through massive protein sequence variation, a property shared uniquely with the adaptive immune response. Little is known about how recognition is achieved by DGR variable proteins. Here, we present the structure of the Bordetella bacteriophage DGR variable protein major tropism determinant (Mtd) bound to the receptor pertactin, revealing remarkable adaptability in the static binding sites of Mtd. Despite large dissimilarities in ligand binding mode, principles underlying selective recognition were strikingly conserved between Mtd and immunoreceptors. Central to this was the differential amplification of binding strengths by avidity (i.e., multivalency), which not only relaxed the demand for optimal complementarity between Mtd and pertactin but also enhanced distinctions among binding events to provide selectivity. A quantitatively similar balance between complementarity and avidity was observed for Bordetella bacteriophage DGR as occurs in the immune system, suggesting that variable repertoires operate under a narrow set of conditions to recognize novel ligands.
Conflict of interest statement
Figures







References
-
- Pancer Z, Cooper MD. The evolution of adaptive immunity. Annu Rev Immunol. 2006;24:497–518. - PubMed
-
- Davis MM, Bjorkman PJ. T-cell antigen receptor genes and T-cell recognition. Nature. 1988;334:395–402. - PubMed
-
- Alder MN, Rogozin IB, Iyer LM, Glazko GV, Cooper MD, et al. Diversity and function of adaptive immune receptors in a jawless vertebrate. Science. 2005;310:1970–1973. - PubMed
-
- Liu M, Deora R, Doulatov SR, Gingery M, Eiserling FA, et al. Reverse transcriptase-mediated tropism switching in Bordetella bacteriophage. Science. 2002;295:2091–2094. - PubMed
-
- Doulatov S, Hodes A, Dai L, Mandhana N, Liu M, et al. Tropism switching in Bordetella bacteriophage defines a family of diversity-generating retroelements. Nature. 2004;431:476–481. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
