

Cellular multitasking: the dual role of human Cu-ATPases in cofactor delivery and intracellular copper balance
- PMID: 18534184
- PMCID: PMC2556376
- DOI: 10.1016/j.abb.2008.05.005
Cellular multitasking: the dual role of human Cu-ATPases in cofactor delivery and intracellular copper balance
Abstract
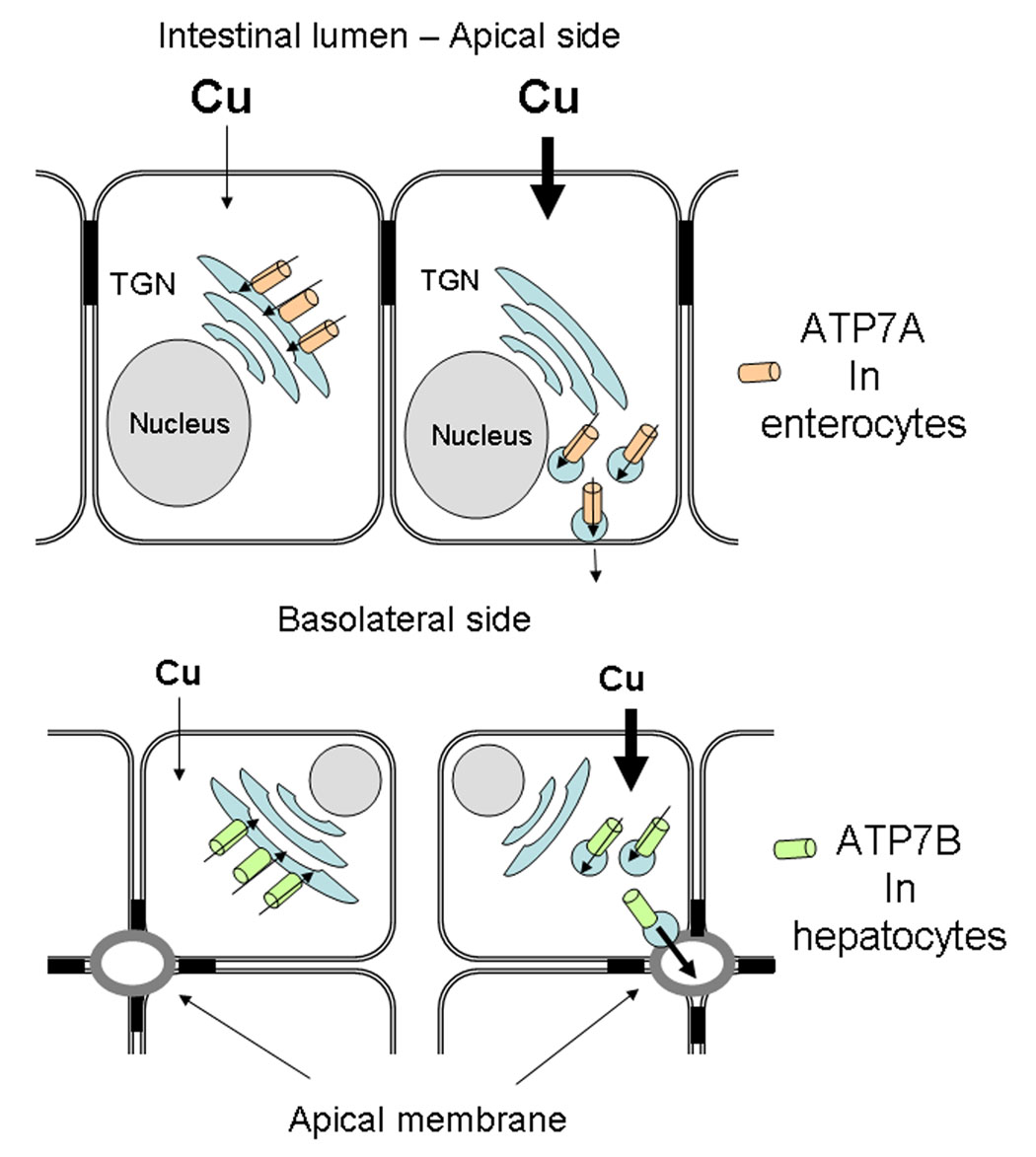
The human copper-transporting ATPases (Cu-ATPases) are essential for dietary copper uptake, normal development and function of the CNS, and regulation of copper homeostasis in the body. In a cell, Cu-ATPases maintain the intracellular concentration of copper by transporting copper into intracellular exocytic vesicles. In addition, these P-type ATPases mediate delivery of copper to copper-dependent enzymes in the secretory pathway and in specialized cell compartments such as secretory granules or melanosomes. The multiple functions of human Cu-ATPase necessitate complex regulation of these transporters that is mediated through the presence of regulatory domains in their structure, posttranslational modification and intracellular trafficking, as well as interactions with the copper chaperone Atox1 and other regulatory molecules. In this review, we summarize the current information on the function and regulatory mechanisms acting on human Cu-ATPases ATP7A and ATP7B. Brief comparison with the Cu-ATPase orthologs from other species is included.
Figures


References
-
- Bartee MY, Lutsenko S. Hepatic copper-transporting ATPase ATP7B: function and inactivation at the molecular and cellular level. Biometals. 2007;20:627–637. - PubMed
-
- Kim BE, Nevitt T, Thiele DJ. Mechanisms for copper acquisition, distribution and regulation. Nat Chem Biol. 2008;4:176–185. - PubMed
-
- Balamurugan K, Schaffner W. Copper homeostasis in eukaryotes: teetering on a tightrope. Biochim Biophys Acta. 2006;1763:737–746. - PubMed
-
- Bertinato J, L'Abbe MR. Maintaining copper homeostasis: regulation of copper-trafficking proteins in response to copper deficiency or overload. J Nutr Biochem. 2004;15:316–322. - PubMed
-
- Field LS, Luk E, Culotta VC. Copper chaperones: personal escorts for metal ions. J Bioenerg Biomembr. 2002;34:373–379. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
