

Hip1r is expressed in gastric parietal cells and is required for tubulovesicle formation and cell survival in mice
- PMID: 18535670
- PMCID: PMC2413185
- DOI: 10.1172/JCI33569
Hip1r is expressed in gastric parietal cells and is required for tubulovesicle formation and cell survival in mice
Abstract
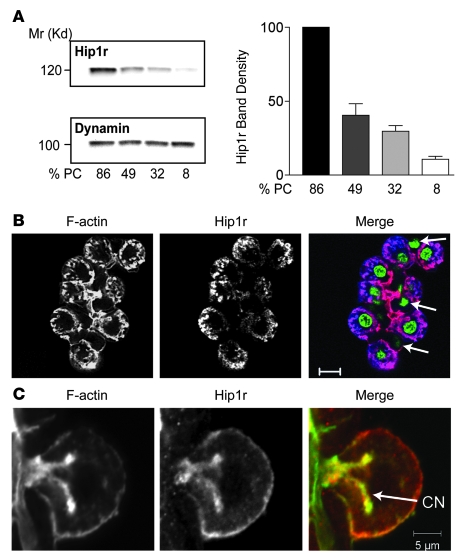
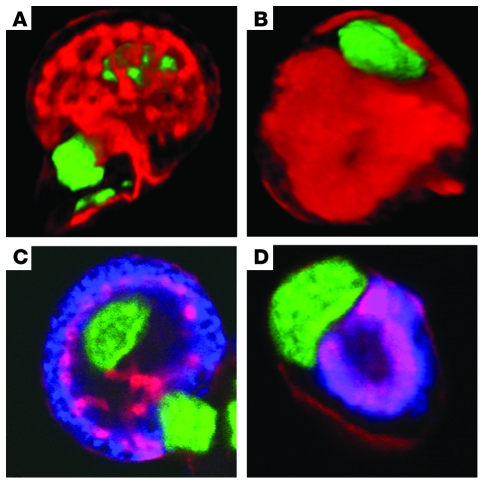
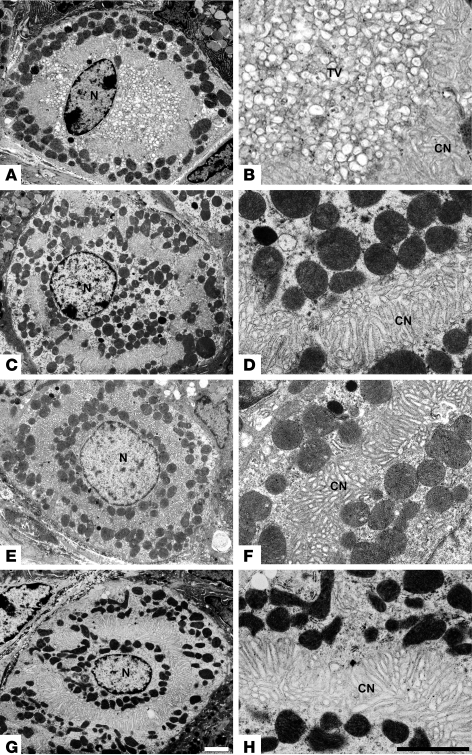
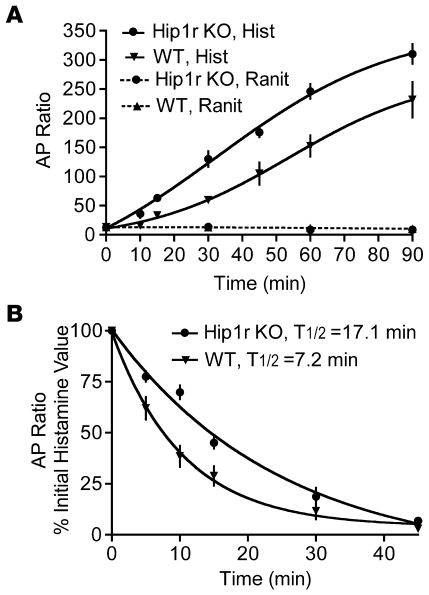
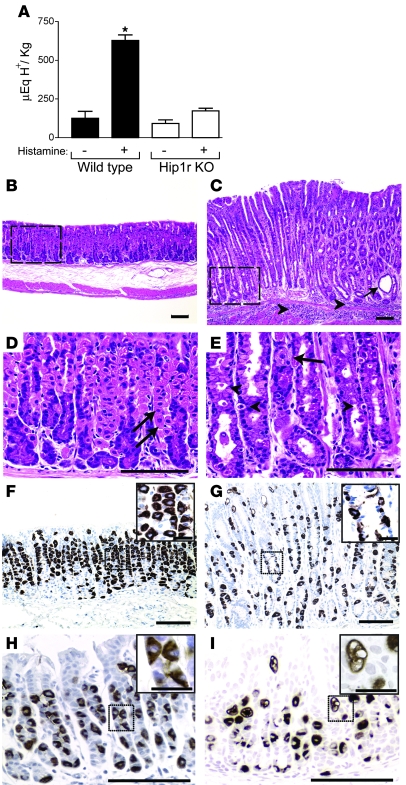
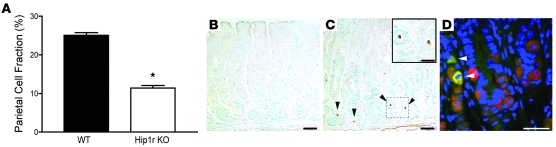
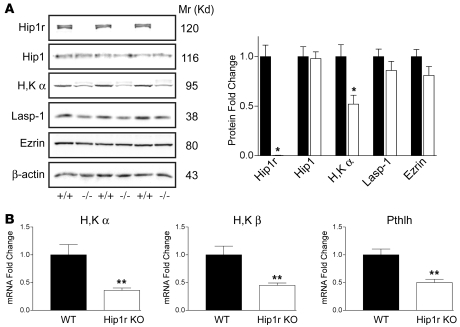
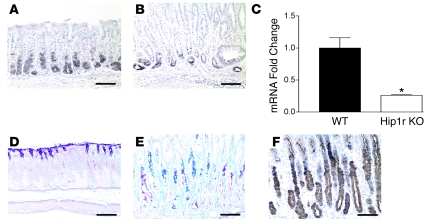
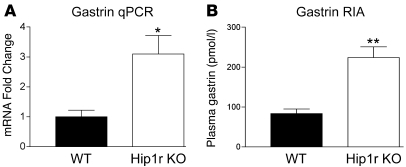
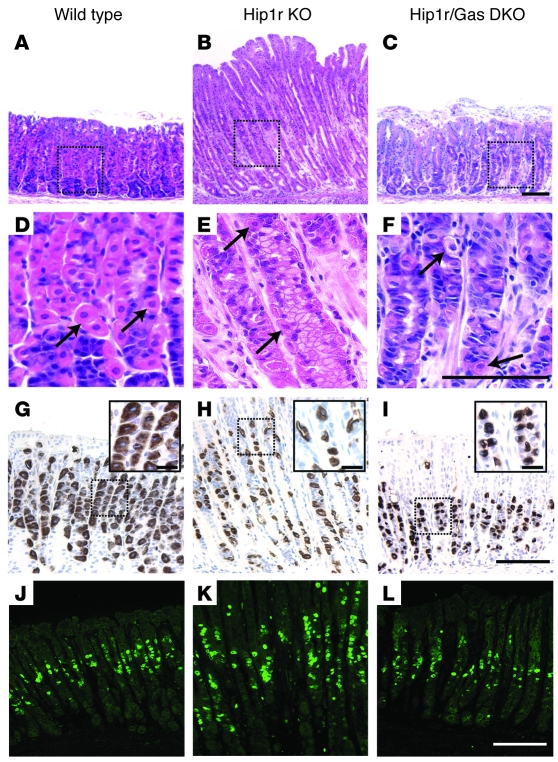
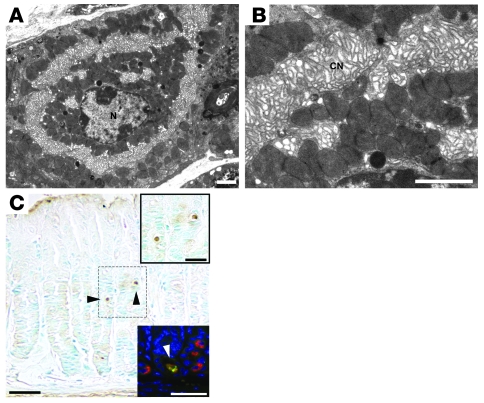
Huntingtin interacting protein 1 related (Hip1r) is an F-actin- and clathrin-binding protein involved in vesicular trafficking. In this study, we demonstrate that Hip1r is abundantly expressed in the gastric parietal cell, predominantly localizing with F-actin to canalicular membranes. Hip1r may provide a critical function in vivo, as demonstrated by extensive changes to parietal cells and the gastric epithelium in Hip1r-deficient mice. Electron microscopy revealed abnormal apical canalicular membranes and loss of tubulovesicles in mutant parietal cells, suggesting that Hip1r is necessary for the normal trafficking of these secretory membranes. Accordingly, acid secretory dynamics were altered in mutant parietal cells, with enhanced activation and acid trapping, as measured in isolated gastric glands. At the whole-organ level, gastric acidity was reduced in Hip1r-deficient mice, and the gastric mucosa was grossly transformed, with fewer parietal cells due to enhanced apoptotic cell death and glandular hypertrophy associated with cellular transformation. Hip1r-deficient mice had increased expression of the gastric growth factor gastrin, and mice mutant for both gastrin and Hip1r exhibited normalization of both proliferation and gland height. Taken together, these studies demonstrate that Hip1r plays a significant role in gastric physiology, mucosal architecture, and secretory membrane dynamics in parietal cells.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
