

A Gata6-Wnt pathway required for epithelial stem cell development and airway regeneration
- PMID: 18536717
- PMCID: PMC2562713
- DOI: 10.1038/ng.157
A Gata6-Wnt pathway required for epithelial stem cell development and airway regeneration
Abstract
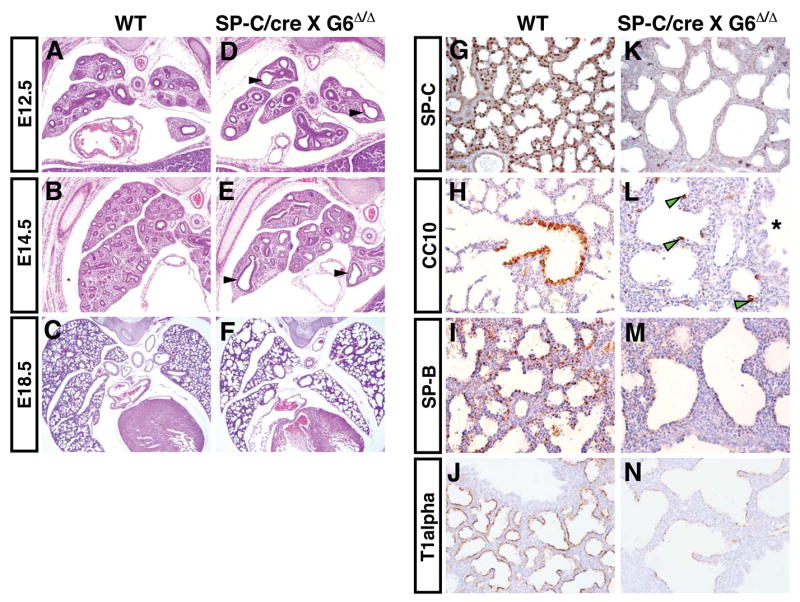
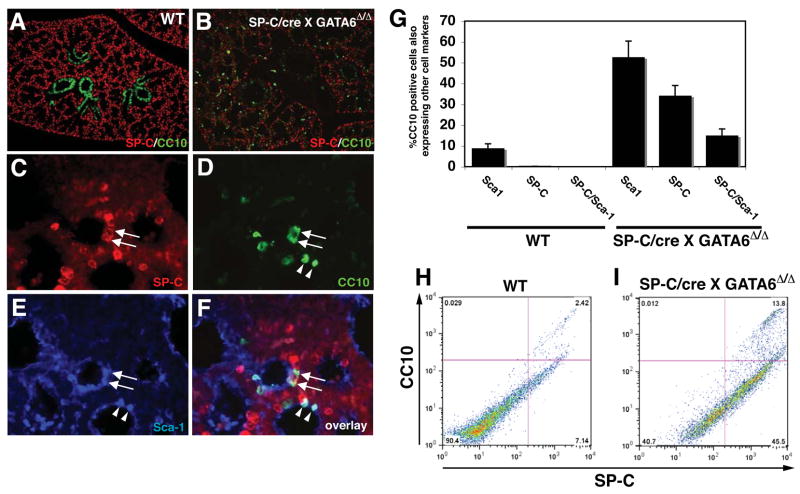
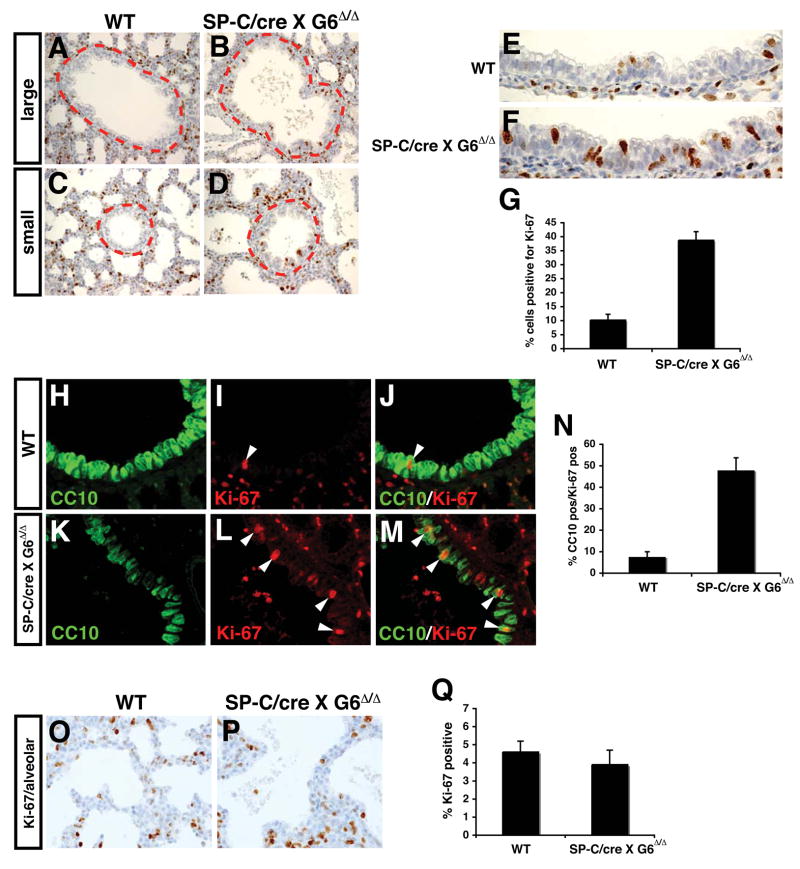
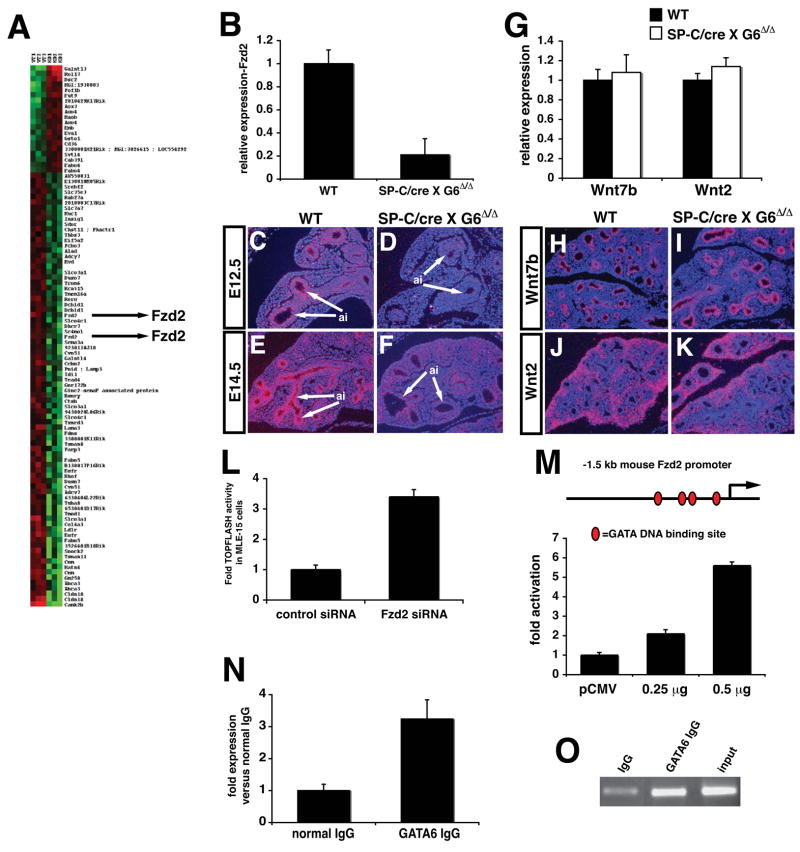
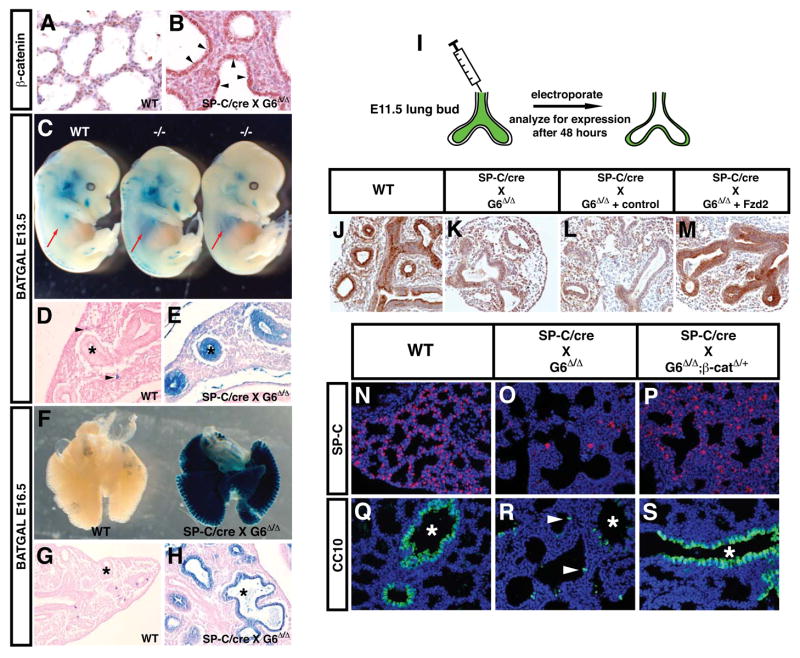
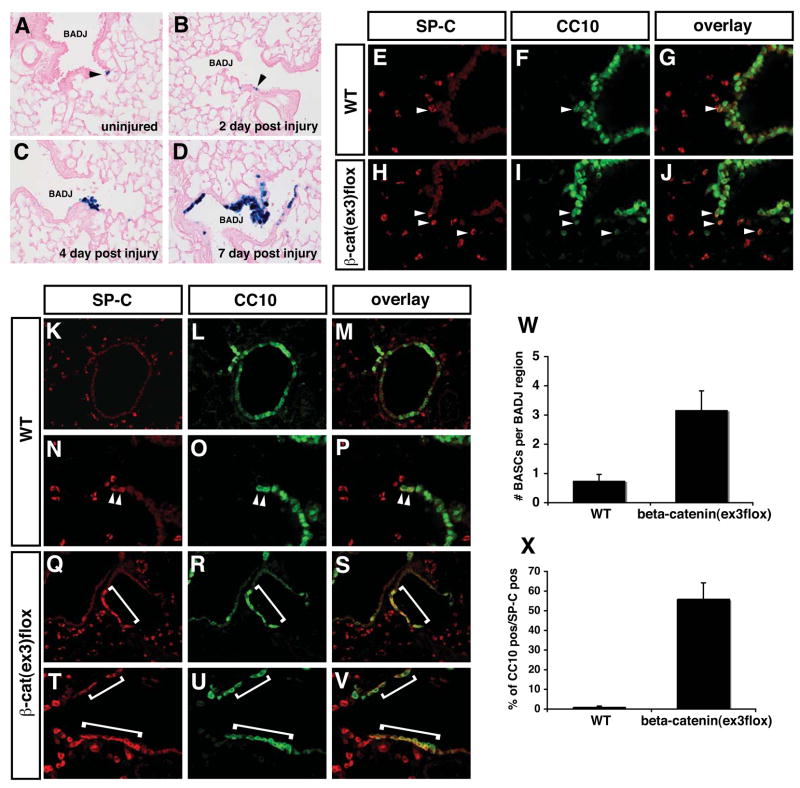
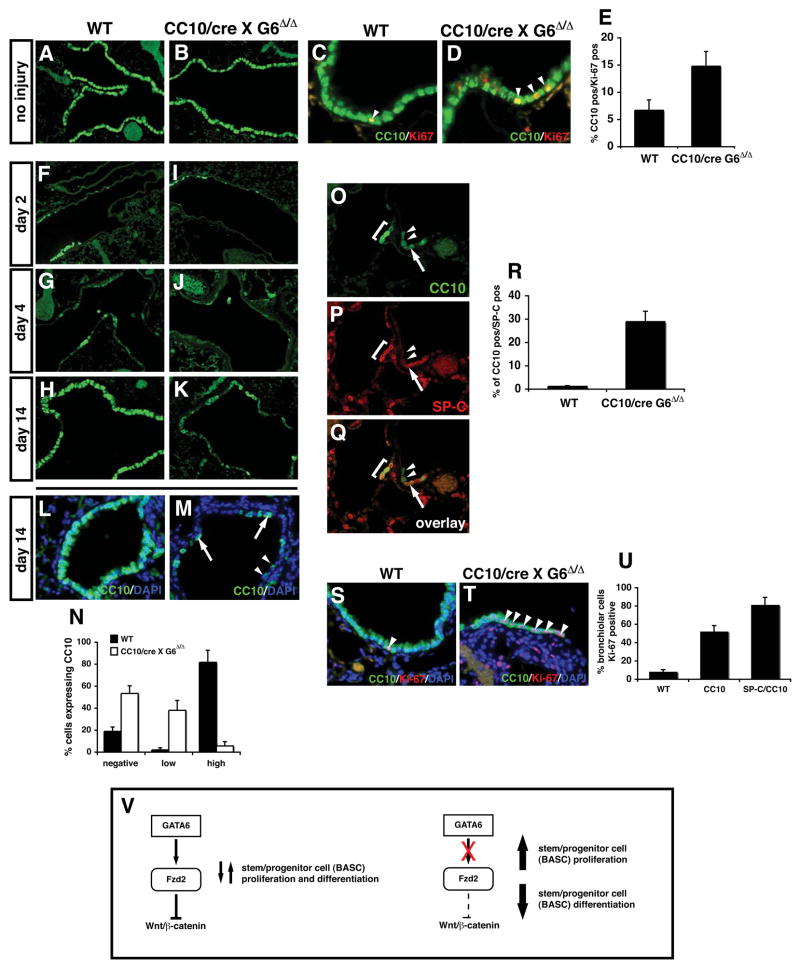
Epithelial organs, including the lung, are known to possess regenerative abilities through activation of endogenous stem cell populations, but the molecular pathways regulating stem cell expansion and regeneration are not well understood. Here we show that Gata6 regulates the temporal appearance and number of bronchioalveolar stem cells (BASCs) in the lung, its absence in Gata6-null lung epithelium leading to the precocious appearance of BASCs and concurrent loss in epithelial differentiation. This expansion of BASCs was the result of a pronounced increase in canonical Wnt signaling in lung epithelium upon loss of Gata6. Expression of the noncanonical Wnt receptor Fzd2 was downregulated in Gata6 mutants and increased Fzd2 or decreased beta-catenin expression rescued, in part, the lung epithelial defects in Gata6 mutants. During lung epithelial regeneration, canonical Wnt signaling was activated in the niche containing BASCs and forced activation of Wnt signaling led to a large increase in BASC numbers. Moreover, Gata6 was required for proper lung epithelial regeneration, and postnatal loss of Gata6 led to increased BASC expansion and decreased differentiation. Together, these data demonstrate that Gata6-regulated Wnt signaling controls the balance between progenitor expansion and epithelial differentiation required for both lung development and regeneration.
Figures
Comment in
-
Lung stem cells in the balance.Nat Genet. 2008 Jul;40(7):822-4. doi: 10.1038/ng0708-822. Nat Genet. 2008. PMID: 18583977 No abstract available.
References
-
- Cardoso WV. Molecular regulation of lung development. Annu Rev Physiol. 2001;63:471–94. - PubMed
-
- Cardoso WV, Lu J. Regulation of early lung morphogenesis: questions, facts and controversies. Development. 2006;133:1611–24. - PubMed
-
- Minoo P, Su G, Drum H, Bringas P, Kimura S. Defects in tracheoesophageal and lung morphogenesis in Nkx2.1 (−/−) mouse embryos. Dev. 1999:60–71. 5/0/209. - PubMed
-
- Wan H, et al. Compensatory roles of Foxa1 and Foxa2 during lungmorphogenesis. J Biol Chem. 2005;280:13809–16. - PubMed
-
- Yang H, Lu MM, Zhang L, Whitsett JA, Morrisey EE. GATA6 regulates differentiation of distal lung epithelium. Development. 2002;129:2233–46. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
