

Differential modulation of AP-1- and CRE-driven transcription by cannabinoid agonists emphasizes functional selectivity at the CB1 receptor
- PMID: 18536748
- PMCID: PMC2527844
- DOI: 10.1038/bjp.2008.230
Differential modulation of AP-1- and CRE-driven transcription by cannabinoid agonists emphasizes functional selectivity at the CB1 receptor
Abstract
Background and purpose: Long-term adaptations to pharmacological stimuli frequently originate from modulation of complex intracellular signalling pathways. We previously reported that HU210 and CP55940, two CB1 cannabinoid receptor agonists, induced opposite effects on TH expression. Herein, we characterized their influence on cAMP response element (CRE) and activator protein 1 (AP-1)-mediated regulation of gene transcription.
Experimental approach: The activity of the agonists was examined on transfected N1E-115 cells in which expression of the luciferase reporter gene was controlled by transcription promoters consisting of repeats of either CRE or AP-1 elements. In addition, the implication of classical signalling pathways was investigated using a variety of kinase inhibitors.
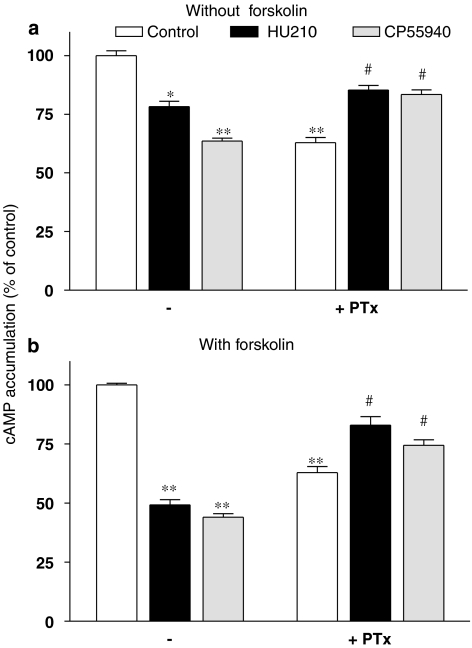
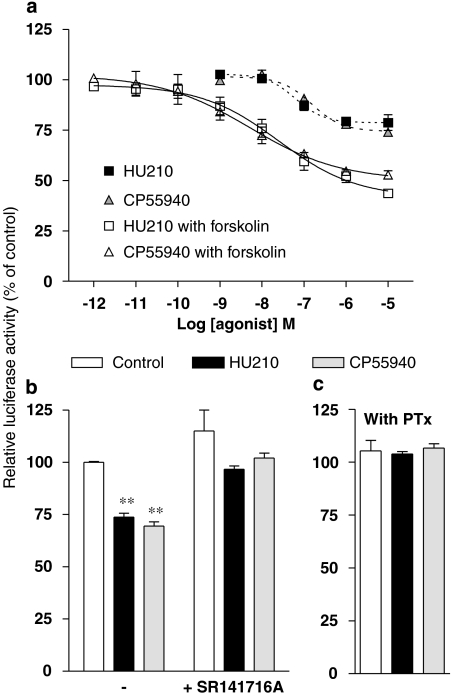
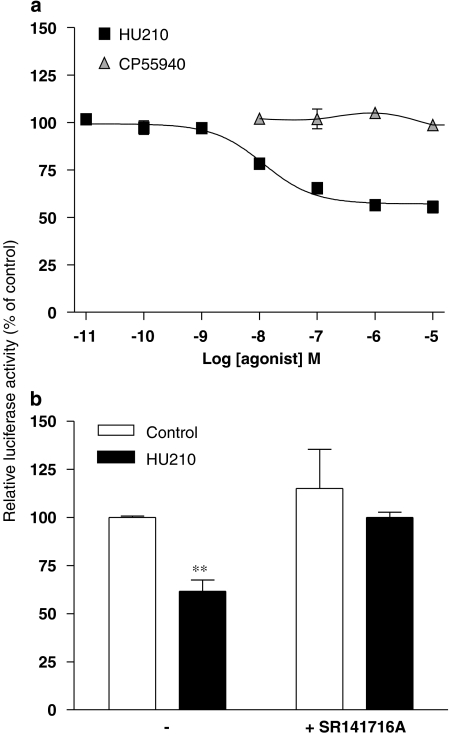
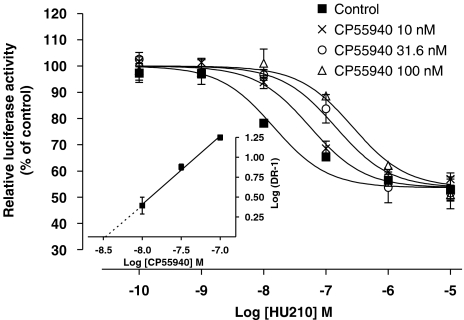
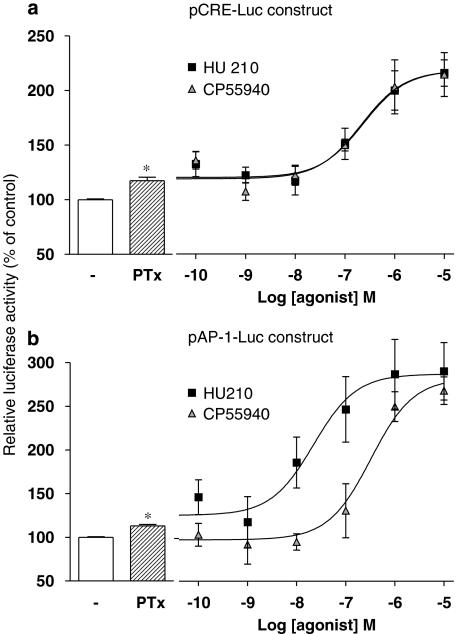
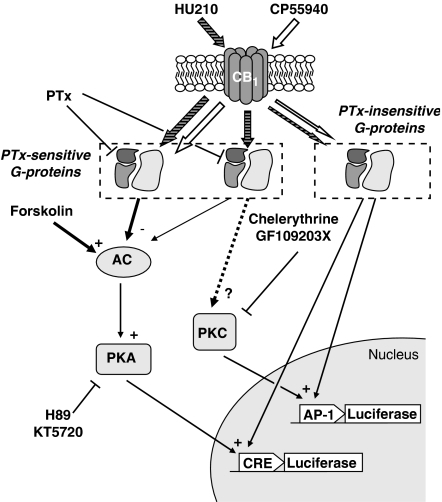
Key results: Consistent with the CB1-mediated reduction of cAMP accumulation, both ligands decreased CRE-driven luciferase expression with similar potencies. HU210 also exhibited a concentration-dependent reduction of luciferase activity in cells engineered to examine AP-1-controlled transcription, whereas such response was not obtained with CP55940. Responses were all inhibited by SR141716A and were modified in Pertussis toxin-treated cells, suggesting agonist-selective regulations of distinct Gi/o-dependent mechanisms through CB1 receptor activation. Finally, PKC inhibitors efficiently inhibited the paradoxical effect of HU210 on AP-1-mediated transcription, indicating selective regulation of PKC-dependent responses.
Conclusions and implications: Together, our results demonstrate that two cannabinoid ligands, commonly used as reference agonists acting on the same receptor with similar affinities, differentially modulate gene transcription through distinct controls of AP-1. This could reflect activation of distinct subsets of Gi/o-proteins, supporting the concept of functional selectivity at CB1 receptors.
Figures
References
-
- Bash R, Rubovitch V, Gafni M, Sarne Y. The stimulatory effect of cannabinoids on calcium uptake is mediated by Gs GTP-binding proteins and cAMP formation. Neurosignals. 2003;12:39–44. - PubMed
-
- Bonhaus DW, Chang LK, Kwan J, Martin GR. Dual activation and inhibition of adenylyl cyclase by cannabinoid receptor agonists: evidence for agonist-specific trafficking of intracellular responses. J Pharmacol Exp Ther. 1998;287:884–888. - PubMed
-
- Bosier B, Hermans E. Versatility of GPCR recognition by drugs: from biological implications to therapeutic relevance. Trends Pharmacol Sci. 2007;28:438–446. - PubMed
-
- Bosier B, Tilleux S, Najimi M, Lambert DM, Hermans E. Agonist selective modulation of tyrosine hydroxylase expression by cannabinoid ligands in a murine neuroblastoma cell line. J Neurochem. 2007;102:1996–2007. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
