

Spontaneous intersubunit rotation in single ribosomes
- PMID: 18538656
- PMCID: PMC2491453
- DOI: 10.1016/j.molcel.2008.05.004
Spontaneous intersubunit rotation in single ribosomes
Abstract
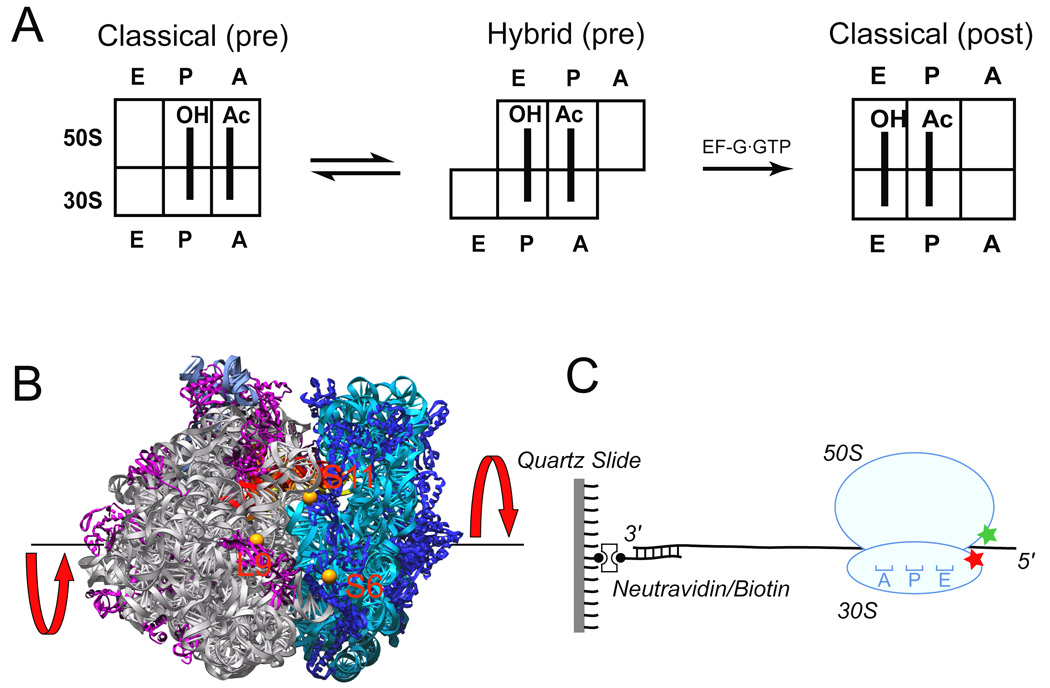
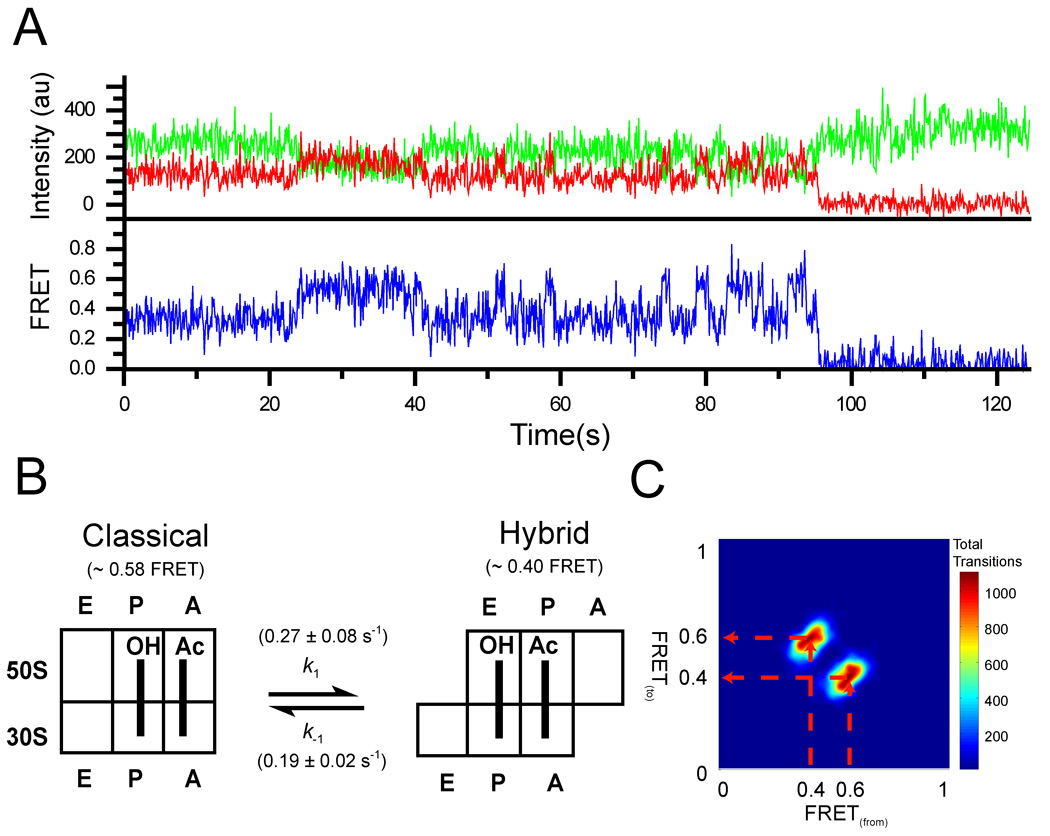
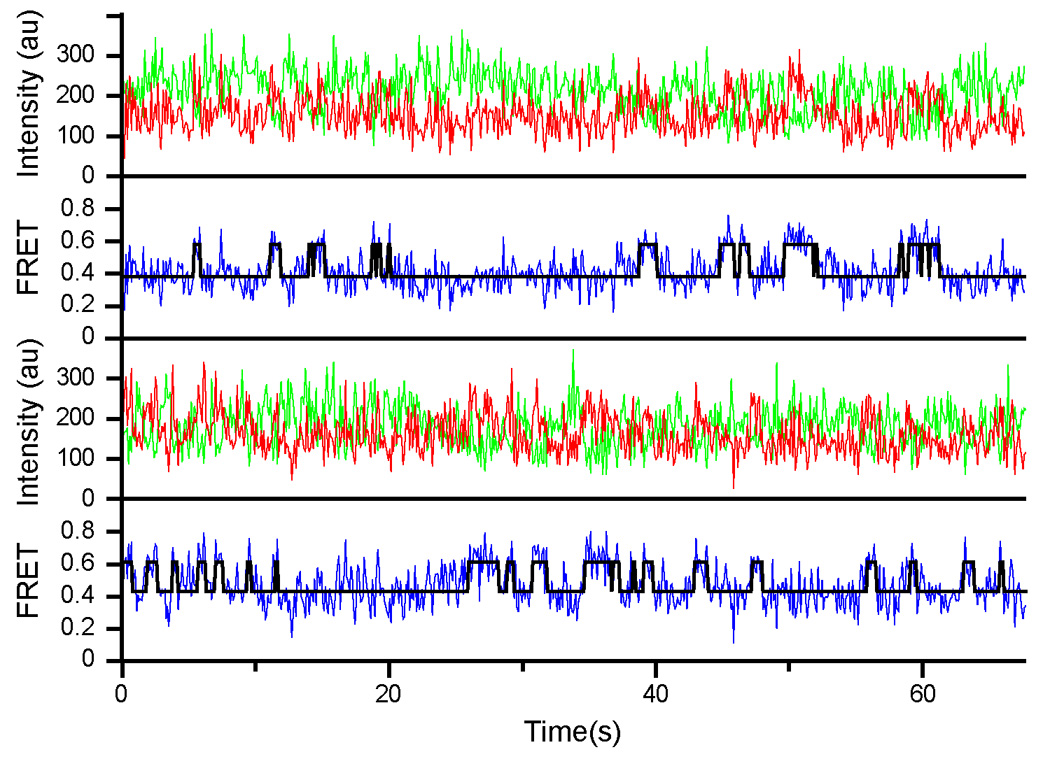
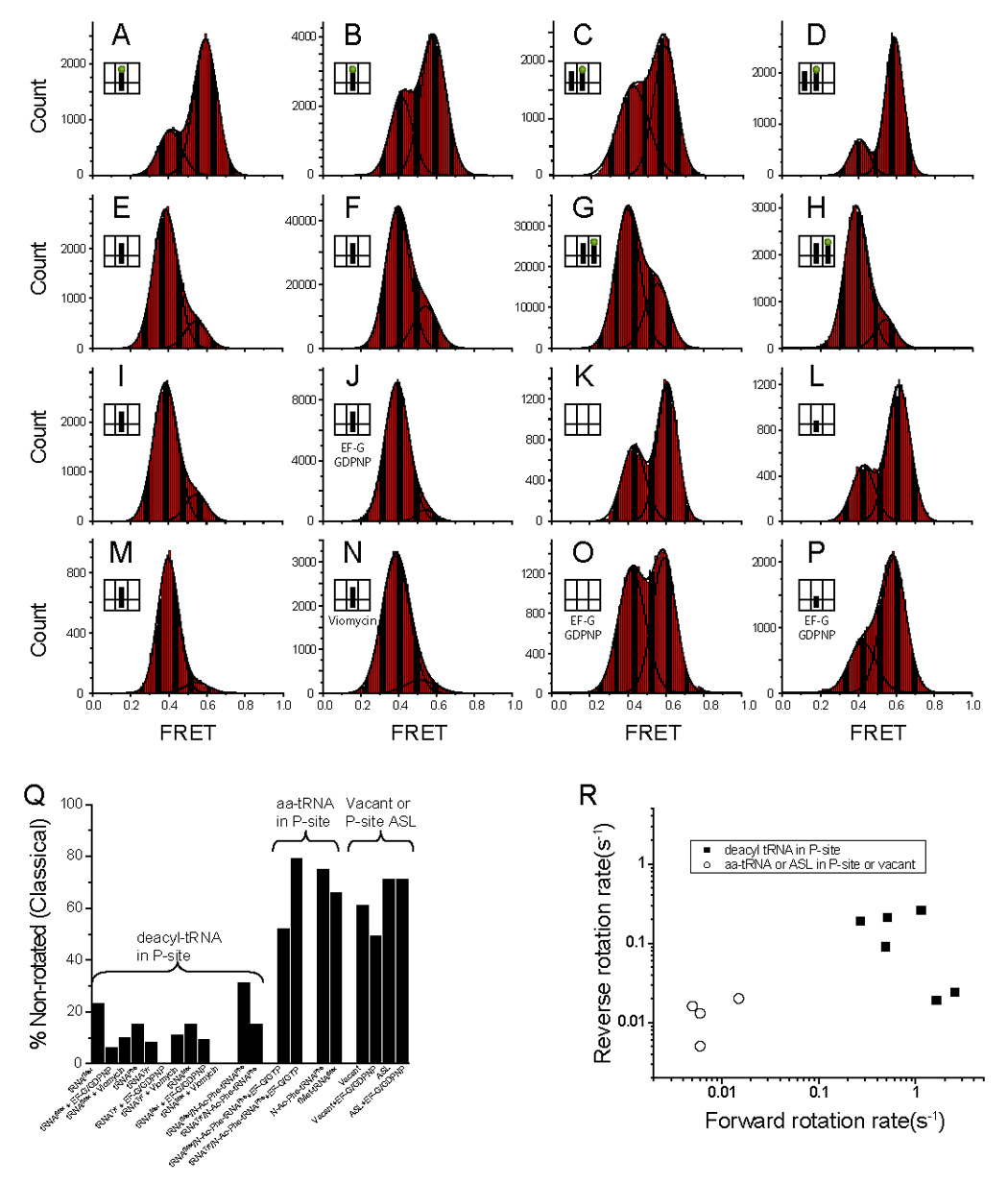
During the elongation cycle, tRNA and mRNA undergo coupled translocation through the ribosome catalyzed by elongation factor G (EF-G). Cryo-EM reconstructions of certain EF-G-containing complexes led to the proposal that the mechanism of translocation involves rotational movement between the two ribosomal subunits. Here, using single-molecule FRET, we observe that pretranslocation ribosomes undergo spontaneous intersubunit rotational movement in the absence of EF-G, fluctuating between two conformations corresponding to the classical and hybrid states of the translocational cycle. In contrast, posttranslocation ribosomes are fixed predominantly in the classical, nonrotated state. Movement of the acceptor stem of deacylated tRNA into the 50S E site and EF-G binding to the ribosome both contribute to stabilization of the rotated, hybrid state. Furthermore, the acylation state of P site tRNA has a dramatic effect on the frequency of intersubunit rotation. Our results provide direct evidence that the intersubunit rotation that underlies ribosomal translocation is thermally driven.
Figures

References
-
- Amand B, Pochon F, Lavalette D. Rotational diffusion of Escherichia coli ribosomes. I. - Free 70 S, 50 S and 30 S particles. Biochimie. 1977;59:779–784. - PubMed
-
- Astumian RD. Thermodynamics and kinetics of a Brownian motor. Science. 1997;276:917–922. - PubMed
-
- Blanchard SC, Gonzalez RL, Kim HD, Chu S, Puglisi JD. tRNA selection and kinetic proofreading in translation. Nat Struct Mol Biol. 2004a;11:1008–1014. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
