

Regulation of cell signaling dynamics by the protein kinase-scaffold Ste5
- PMID: 18538663
- PMCID: PMC2518723
- DOI: 10.1016/j.molcel.2008.04.016
Regulation of cell signaling dynamics by the protein kinase-scaffold Ste5
Abstract
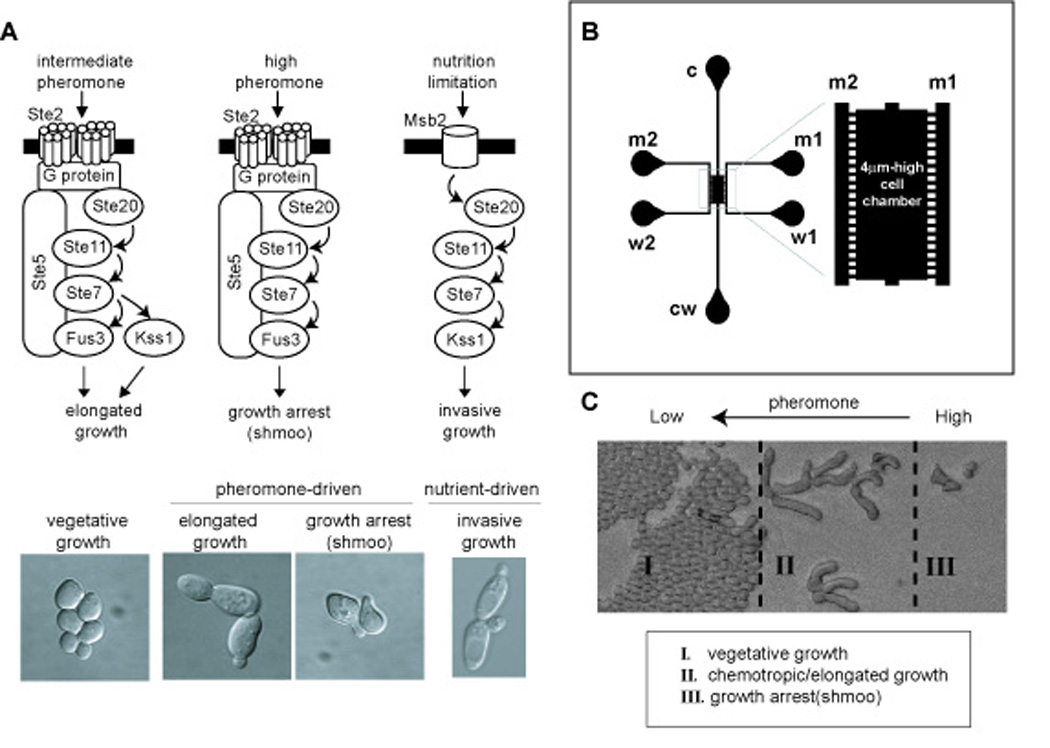
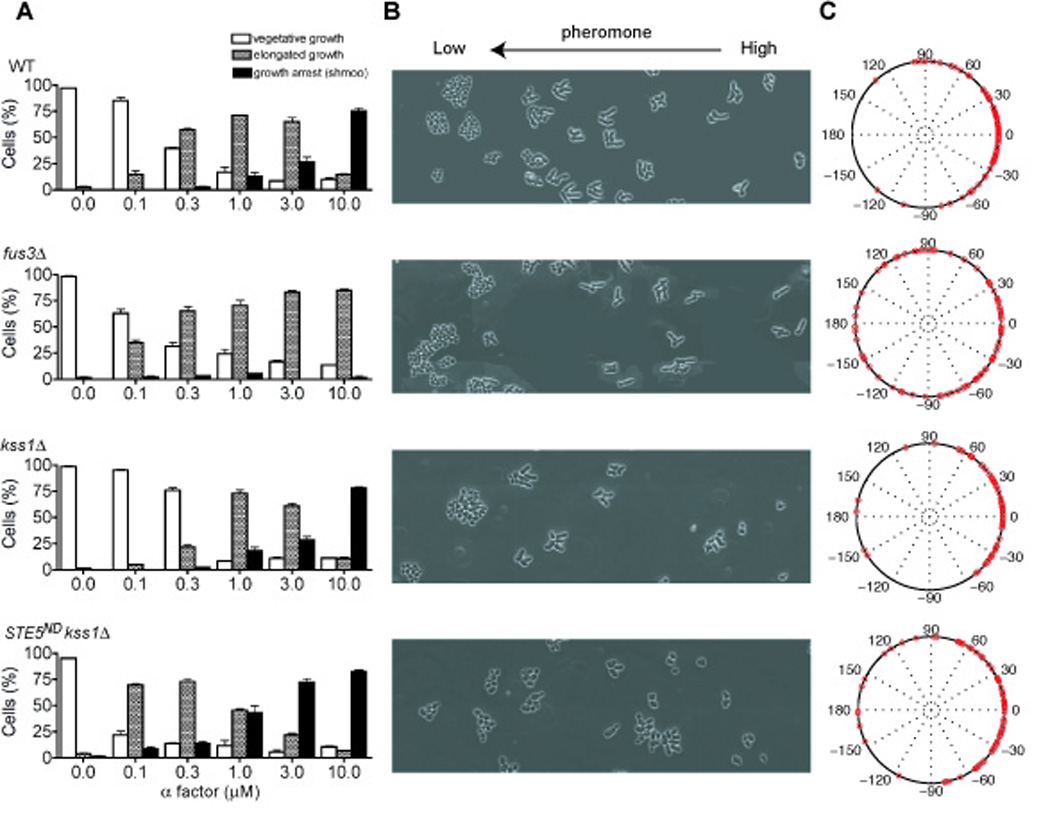
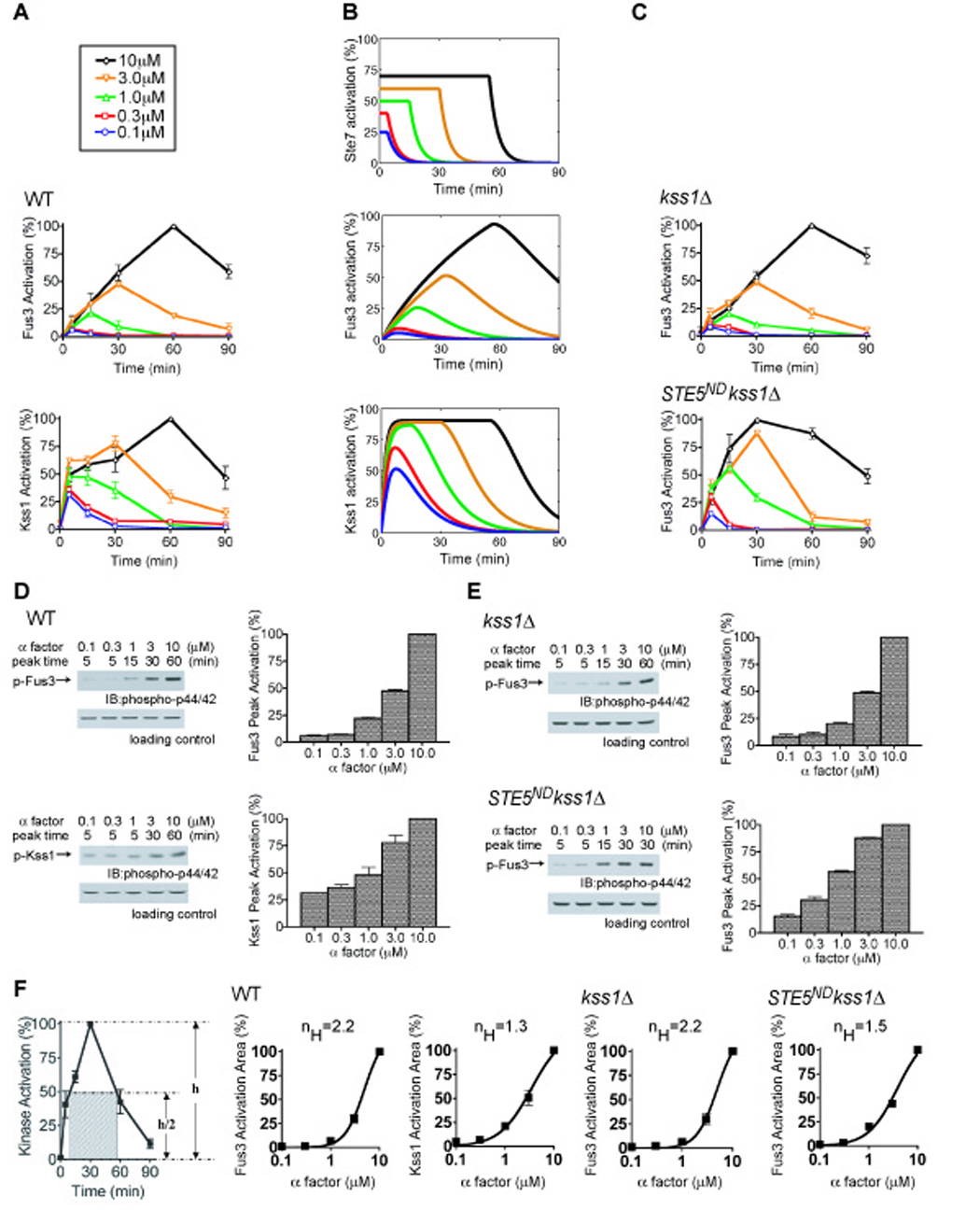
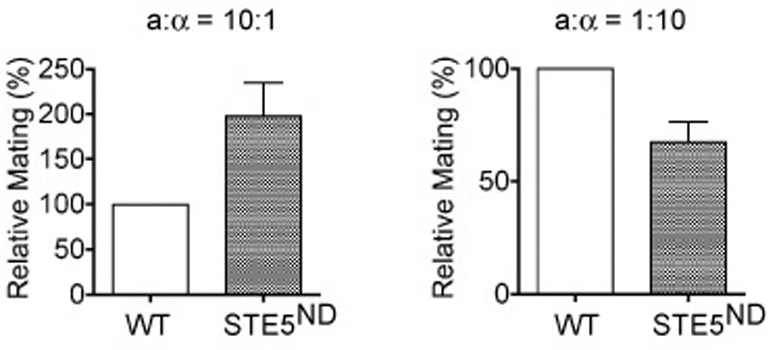
Cell differentiation requires the ability to detect and respond appropriately to a variety of extracellular signals. Here we investigate a differentiation switch induced by changes in the concentration of a single stimulus. Yeast cells exposed to high doses of mating pheromone undergo cell division arrest. Cells at intermediate doses become elongated and divide in the direction of a pheromone gradient (chemotropic growth). Either of the pheromone-responsive MAP kinases, Fus3 and Kss1, promotes cell elongation, but only Fus3 promotes chemotropic growth. Whereas Kss1 is activated rapidly and with a graded dose-response profile, Fus3 is activated slowly and exhibits a steeper dose-response relationship (ultrasensitivity). Fus3 activity requires the scaffold protein Ste5; when binding to Ste5 is abrogated, Fus3 behaves like Kss1, and the cells no longer respond to a gradient or mate efficiently with distant partners. We propose that scaffold proteins serve to modulate the temporal and dose-response behavior of the MAP kinase.
Figures
References
-
- Bhattacharyya RP, Remenyi A, Good MC, Bashor CJ, Falick AM, Lim WA. The Ste5 scaffold allosterically modulates signaling output of the yeast mating pathway. Science. 2006;311:822–826. - PubMed
-
- Breitkreutz A, Boucher L, Tyers M. MAPK specificity in the yeast pheromone response independent of transcriptional activation. Curr Biol. 2001;11:1266–1271. - PubMed
-
- Choi KY, Satterberg B, Lyons DM, Elion EA. Ste5 tethers multiple protein kinases in the MAP kinase cascade required for mating in S. cerevisiae. Cell. 1994;78:499–512. - PubMed
-
- Cook JG, Bardwell L, Thorner J. Inhibitory and activating functions for MAPK Kss1 in the S. cerevisiae filamentous-growth signalling pathway. Nature. 1997;390:85–88. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
