

Involvement of the MADS-box gene ZMM4 in floral induction and inflorescence development in maize
- PMID: 18539775
- PMCID: PMC2492622
- DOI: 10.1104/pp.107.115261
Involvement of the MADS-box gene ZMM4 in floral induction and inflorescence development in maize
Abstract
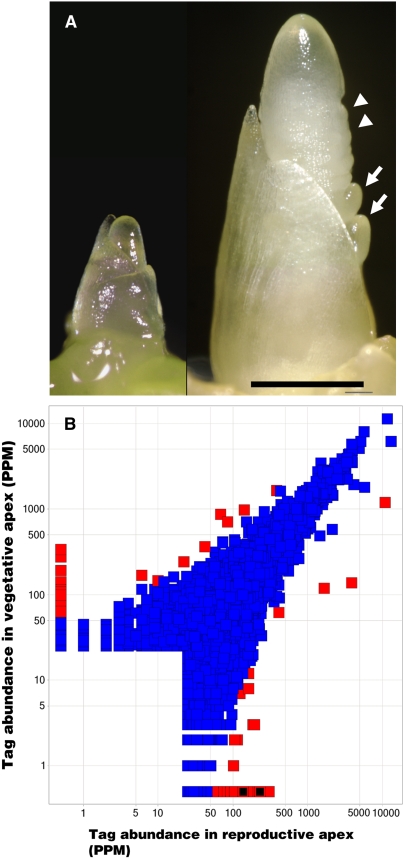
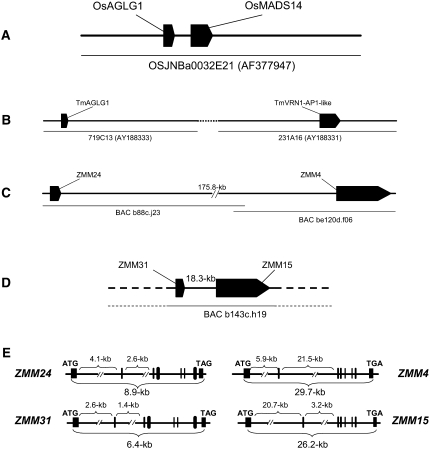
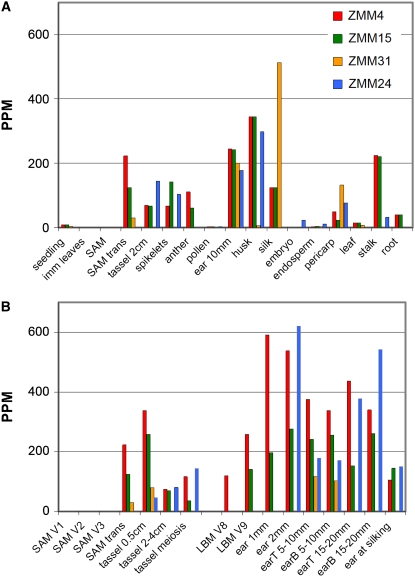
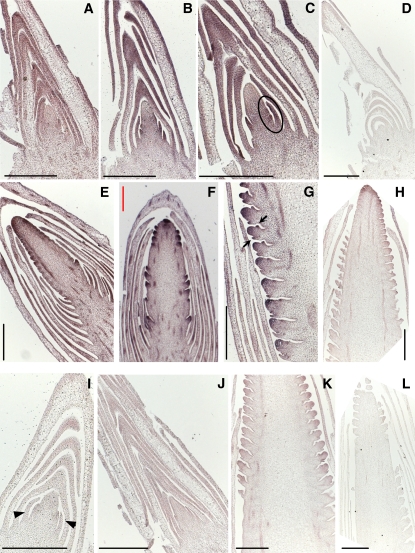
The switch from vegetative to reproductive growth is marked by the termination of vegetative development and the adoption of floral identity by the shoot apical meristem (SAM). This process is called the floral transition. To elucidate the molecular determinants involved in this process, we performed genome-wide RNA expression profiling on maize (Zea mays) shoot apices at vegetative and early reproductive stages using massively parallel signature sequencing technology. Profiling revealed significant up-regulation of two maize MADS-box (ZMM) genes, ZMM4 and ZMM15, after the floral transition. ZMM4 and ZMM15 map to duplicated regions on chromosomes 1 and 5 and are linked to neighboring MADS-box genes ZMM24 and ZMM31, respectively. This gene order is syntenic with the vernalization1 locus responsible for floral induction in winter wheat (Triticum monococcum) and similar loci in other cereals. Analyses of temporal and spatial expression patterns indicated that the duplicated pairs ZMM4-ZMM24 and ZMM15-ZMM31 are coordinately activated after the floral transition in early developing inflorescences. More detailed analyses revealed ZMM4 expression initiates in leaf primordia of vegetative shoot apices and later increases within elongating meristems acquiring inflorescence identity. Expression analysis in late flowering mutants positioned all four genes downstream of the floral activators indeterminate1 (id1) and delayed flowering1 (dlf1). Overexpression of ZMM4 leads to early flowering in transgenic maize and suppresses the late flowering phenotype of both the id1 and dlf1 mutations. Our results suggest ZMM4 may play roles in both floral induction and inflorescence development.
Figures
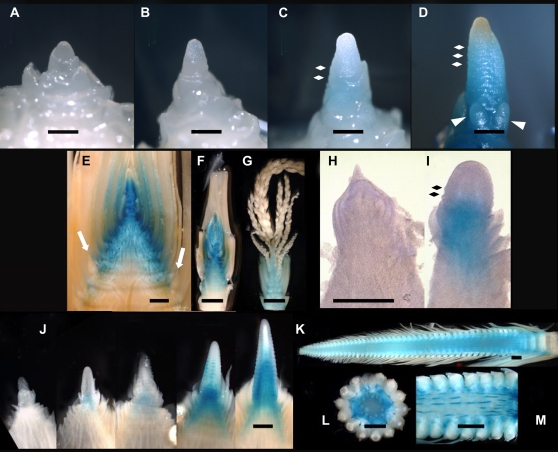
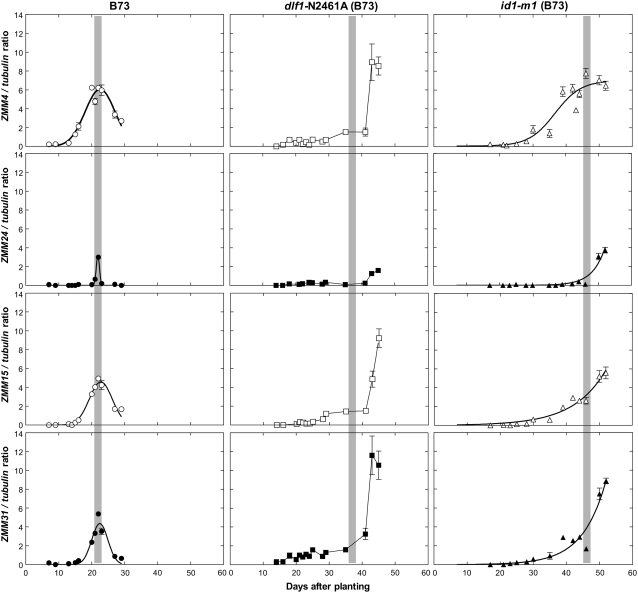
References
-
- Abe M, Kobayashi Y, Yamamoto S, Daimon Y, Yamaguchi A, Ikeda Y, Ichinoki H, Notaguchi M, Goto K, Araki T (2005) FD, a bZIP protein mediating signals from the floral pathway integrator FT at the shoot apex. Science 309 1052–1056 - PubMed
-
- Becker A, Theissen G (2003) The major clades of MADS-box genes and their role in the development and evolution of flowering plants. Mol Phylogenet Evol 29 464–489 - PubMed
-
- Becker A, Winter KU, Meyer B, Saedler H, Theissen G (2000) MADS-Box gene diversity in seed plants 300 million years ago. Mol Biol Evol 17 1425–1434 - PubMed
-
- Bernier G, Perilleux C (2005) A physiological overview of the genetics of flowering time control. Plant Biotechnol J 3 3–16 - PubMed
-
- Blazquez MA, Soowal LN, Lee I, Weigel D (1997) LEAFY expression and flower initiation in Arabidopsis. Development 124 3835–3844 - PubMed
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
