

Glutamate forward and reverse transport: from molecular mechanism to transporter-mediated release after ischemia
- PMID: 18543277
- PMCID: PMC2632779
- DOI: 10.1002/iub.98
Glutamate forward and reverse transport: from molecular mechanism to transporter-mediated release after ischemia
Abstract
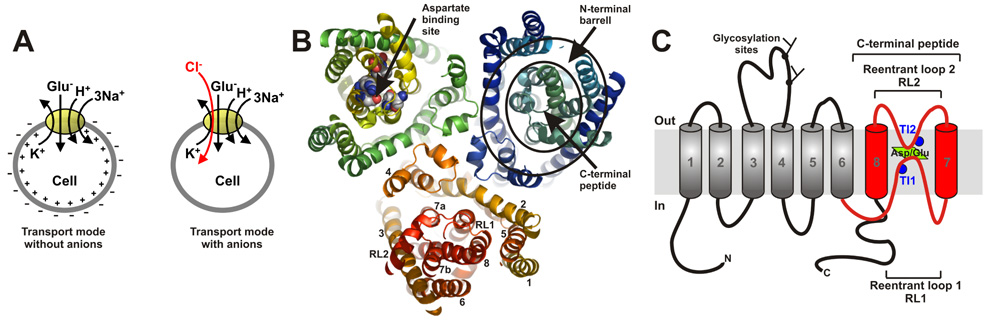
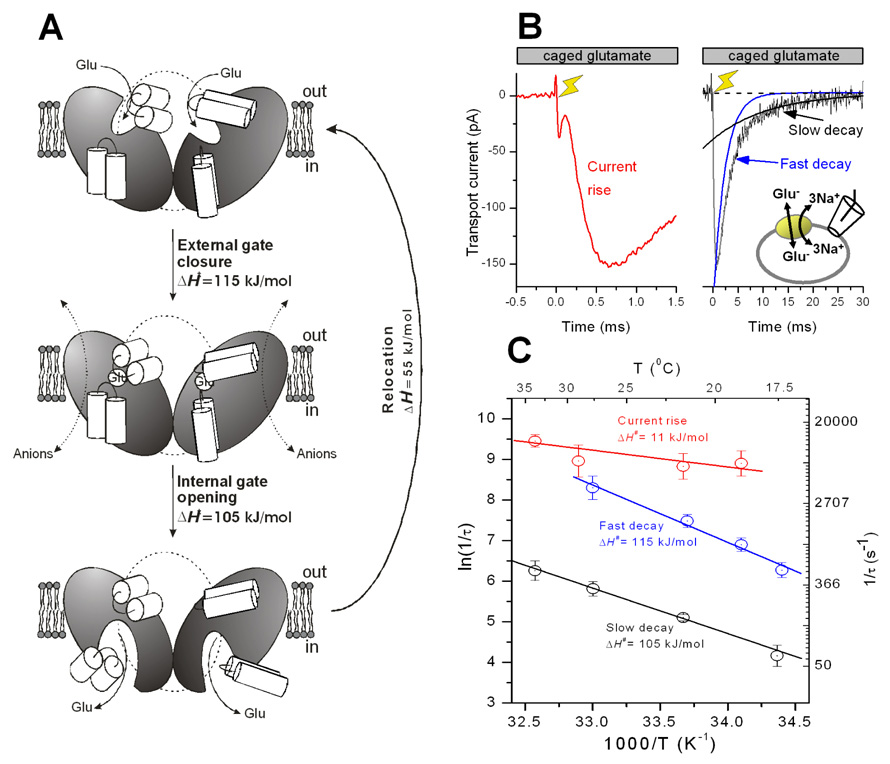
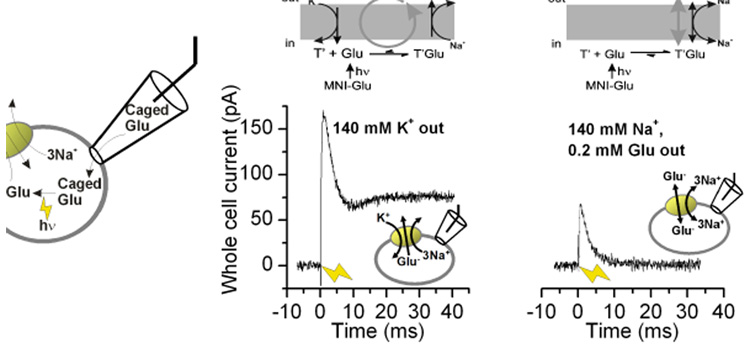
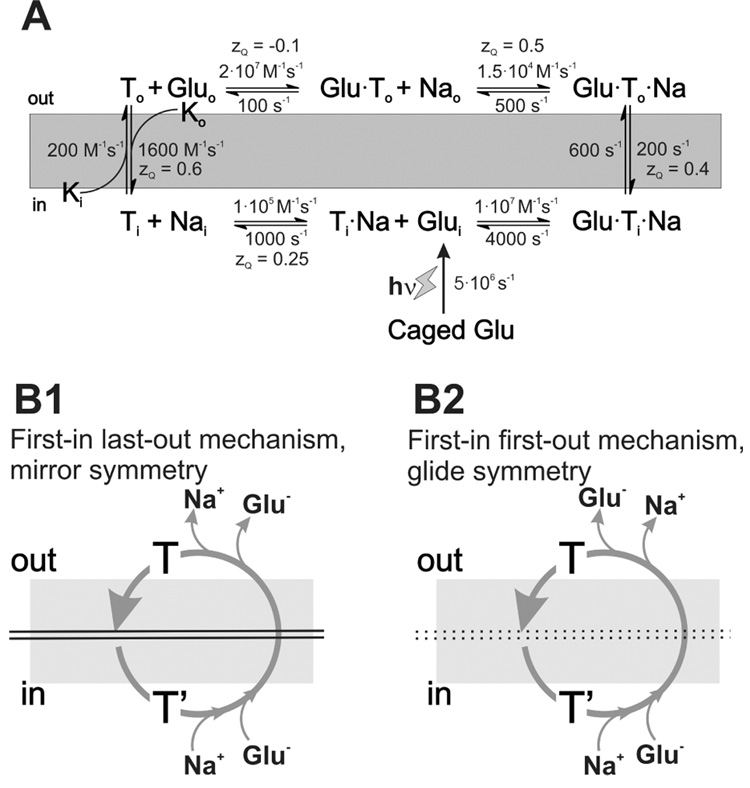
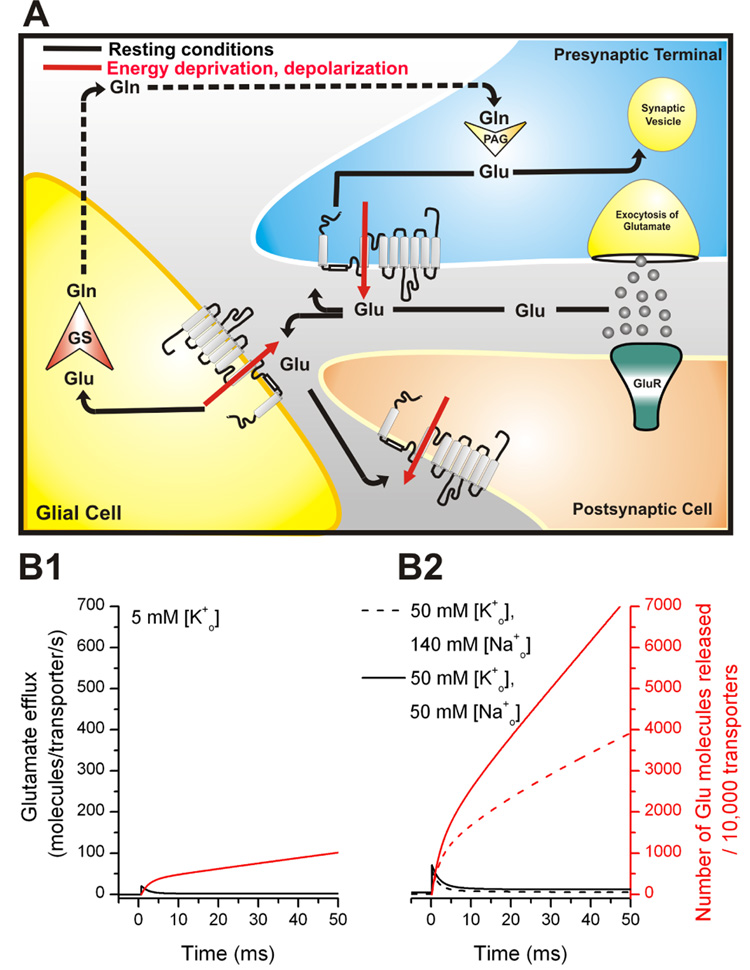
Glutamate transporters remove the excitatory neurotransmitter glutamate from the extracellular space after neurotransmission is complete, by taking glutamate up into neurons and glia cells. As thermodynamic machines, these transporters can also run in reverse, releasing glutamate into the extracellular space. Because glutamate is excitotoxic, this transporter-mediated release is detrimental to the health of neurons and axons, and it, thus, contributes to the brain damage that typically follows a stroke. This review highlights current ideas about the molecular mechanisms underlying glutamate uptake and glutamate reverse transport. It also discusses the implications of transporter-mediated glutamate release for cellular function under physiological and patho-physiological conditions.
Copyright 2008 IUBMB
Figures
References
-
- Danbolt NC. Glutamate uptake. Prog. Neurobiol. 2001;65:101–105. - PubMed
-
- Vandenberg RJ. Molecular pharmacology and physiology of glutamate transporters in the central nervous system. Clin. Exp. Pharmacol. Physiol. 1998;25:393–400. - PubMed
-
- Kanai Y, Hediger MA. The glutamate and neutral amino acid transporter family: physiological and pharmacological implications. Eur. J. Pharmacol. 2003;479:237–247. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
