

X-ray structure of ILL2, an auxin-conjugate amidohydrolase from Arabidopsis thaliana
- PMID: 18543330
- PMCID: PMC2605170
- DOI: 10.1002/prot.22124
X-ray structure of ILL2, an auxin-conjugate amidohydrolase from Arabidopsis thaliana
Abstract
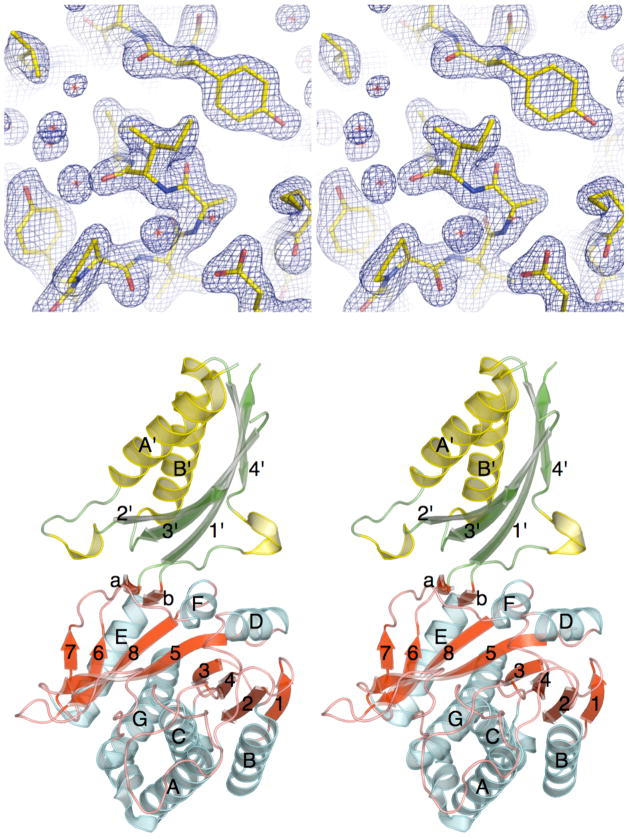
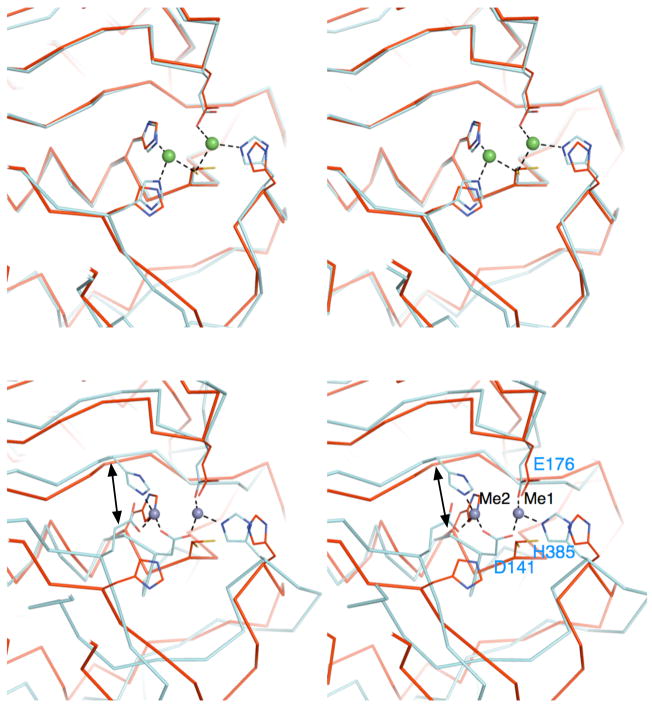
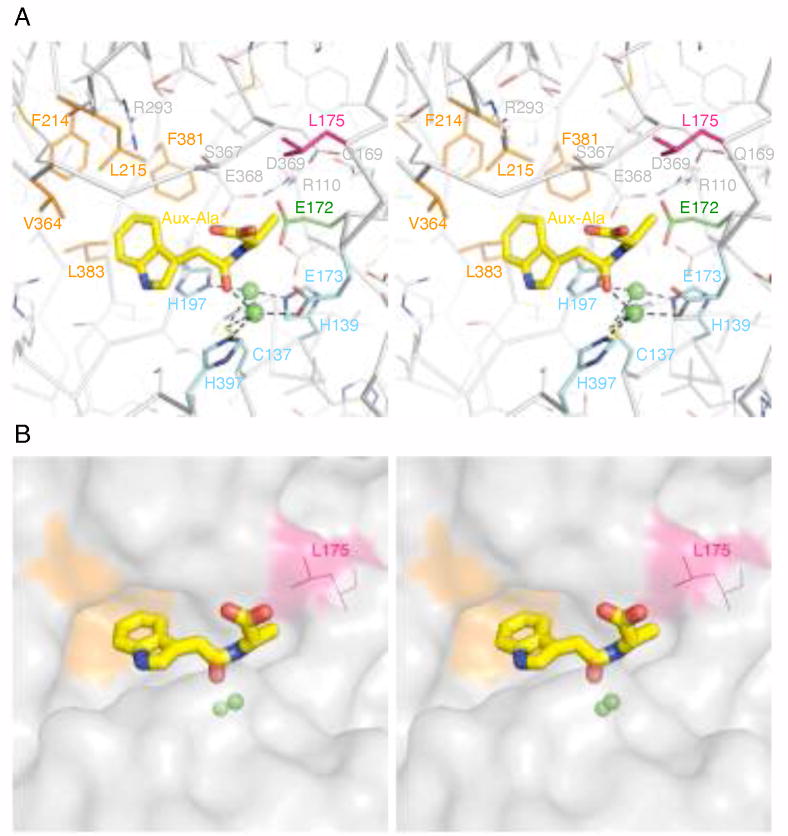
The plant hormone indole-3-acetic acid (IAA) is the most abundant natural auxin involved in many aspects of plant development and growth. The IAA levels in plants are modulated by a specific group of amidohydrolases from the peptidase M20D family that release the active hormone from its conjugated storage forms. Here, we describe the X-ray crystal structure of IAA-amino acid hydrolase IAA-leucine resistantlike gene 2 (ILL2) from Arabidopsis thaliana at 2.0 A resolution. ILL2 preferentially hydrolyses the auxin-amino acid conjugate N-(indol-3-acetyl)-alanine. The overall structure of ILL2 is reminiscent of dinuclear metallopeptidases from the M20 peptidase family. The structure consists of two domains, a larger catalytic domain with three-layer alpha beta alpha sandwich architecture and aminopeptidase topology and a smaller satellite domain with two-layer alphabeta-sandwich architecture and alpha-beta-plaits topology. The metal-coordinating residues in the active site of ILL2 include a conserved cysteine that clearly distinguishes this protein from previously structurally characterized members of the M20 peptidase family. Modeling of N-(indol-3-acetyl)-alanine into the active site of ILL2 suggests that Leu175 serves as a key determinant for the amino acid side-chain specificity of this enzyme. Furthermore, a hydrophobic pocket nearby the catalytic dimetal center likely recognizes the indolyl moiety of the substrate. Finally, the active site of ILL2 harbors an absolutely conserved glutamate (Glu172), which is well positioned to act as a general acid-base residue. Overall, the structure of ILL2 suggests that this enzyme likely uses a catalytic mechanism that follows the paradigm established for the other enzymes of the M20 peptidase family.
(c) 2008 Wiley-Liss, Inc.
Figures

Similar articles
-
Hydrolases of the ILR1-like family of Arabidopsis thaliana modulate auxin response by regulating auxin homeostasis in the endoplasmic reticulum.Sci Rep. 2016 Apr 11;6:24212. doi: 10.1038/srep24212. Sci Rep. 2016. PMID: 27063913 Free PMC article.
-
A family of auxin-conjugate hydrolases that contributes to free indole-3-acetic acid levels during Arabidopsis germination.Plant Physiol. 2004 Jun;135(2):978-88. doi: 10.1104/pp.104.039677. Epub 2004 May 21. Plant Physiol. 2004. PMID: 15155875 Free PMC article.
-
Characterization of a family of IAA-amino acid conjugate hydrolases from Arabidopsis.J Biol Chem. 2002 Jun 7;277(23):20446-52. doi: 10.1074/jbc.M111955200. Epub 2002 Mar 28. J Biol Chem. 2002. PMID: 11923288
-
Crystal structure of the indole-3-acetic acid-catabolizing enzyme DAO1 from Arabidopsis thaliana.J Struct Biol. 2020 Dec 1;212(3):107632. doi: 10.1016/j.jsb.2020.107632. Epub 2020 Sep 24. J Struct Biol. 2020. PMID: 32980521
-
The AMI1 gene family: indole-3-acetamide hydrolase functions in auxin biosynthesis in plants.J Exp Bot. 2010;61(1):25-32. doi: 10.1093/jxb/erp292. J Exp Bot. 2010. PMID: 19887500 Review.
Cited by
-
Structural and functional analyses reveal that Staphylococcus aureus antibiotic resistance factor HmrA is a zinc-dependent endopeptidase.J Biol Chem. 2011 Jul 22;286(29):25697-709. doi: 10.1074/jbc.M111.247437. Epub 2011 May 27. J Biol Chem. 2011. PMID: 21622555 Free PMC article.
-
THOUSAND-GRAIN WEIGHT 6, which is an IAA-glucose hydrolase, preferentially recognizes the structure of the indole ring.Sci Rep. 2024 Mar 21;14(1):6778. doi: 10.1038/s41598-024-57506-z. Sci Rep. 2024. PMID: 38514802 Free PMC article.
-
Mutational and structural analysis of L-N-carbamoylase reveals new insights into a peptidase M20/M25/M40 family member.J Bacteriol. 2012 Nov;194(21):5759-68. doi: 10.1128/JB.01056-12. Epub 2012 Aug 17. J Bacteriol. 2012. PMID: 22904279 Free PMC article.
-
Structure of an amidohydrolase, SACOL0085, from methicillin-resistant Staphylococcus aureus COL.Acta Crystallogr Sect F Struct Biol Cryst Commun. 2013 Feb 1;69(Pt 2):103-8. doi: 10.1107/S1744309112049822. Epub 2013 Jan 30. Acta Crystallogr Sect F Struct Biol Cryst Commun. 2013. PMID: 23385746 Free PMC article.
-
Indole-3-acetylaspartate and indole-3-acetylglutamate, the IAA-amide conjugates in the diploid strawberry achene, are hydrolyzed in growing seedlings.Planta. 2019 Apr;249(4):1073-1085. doi: 10.1007/s00425-018-3061-0. Epub 2018 Dec 8. Planta. 2019. PMID: 30535588
References
-
- Bartel B. Auxin Biosynthesis. Annu Rev Plant Physiol Plant Mol Biol. 1997;48:51–66. - PubMed
-
- Ljung K, Hull AK, Kowalczyk M, Marchant A, Celenza J, Cohen JD, Sandberg G. Biosynthesis, conjugation, catabolism and homeostasis of indole-3-acetic acid in Arabidopsis thaliana. Plant Mol Biol. 2002;49(3–4):249–272. - PubMed
-
- Tan X, Calderon-Villalobos LI, Sharon M, Zheng C, Robinson CV, Estelle M, Zheng N. Mechanism of auxin perception by the TIR1 ubiquitin ligase. Nature. 2007;446(7136):640–645. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
