

New insights into mechanism and regulation of actin capping protein
- PMID: 18544499
- PMCID: PMC2583073
- DOI: 10.1016/S1937-6448(08)00604-7
New insights into mechanism and regulation of actin capping protein
Abstract
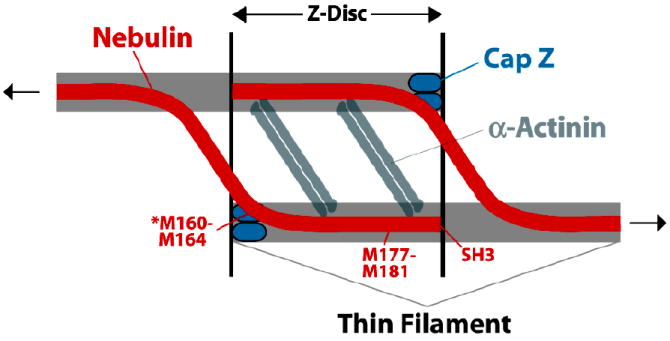
The heterodimeric actin capping protein, referred to here as "CP," is an essential element of the actin cytoskeleton, binding to the barbed ends of actin filaments and regulating their polymerization. In vitro, CP has a critical role in the dendritic nucleation process of actin assembly mediated by Arp2/3 complex, and in vivo, CP is important for actin assembly and actin-based process of morphogenesis and differentiation. Recent studies have provided new insight into the mechanism of CP binding the barbed end, which raises new possibilities for the dynamics of CP and actin in cells. In addition, a number of molecules that bind and regulate CP have been discovered, suggesting new ideas for how CP may integrate into diverse processes of cell physiology.
Figures
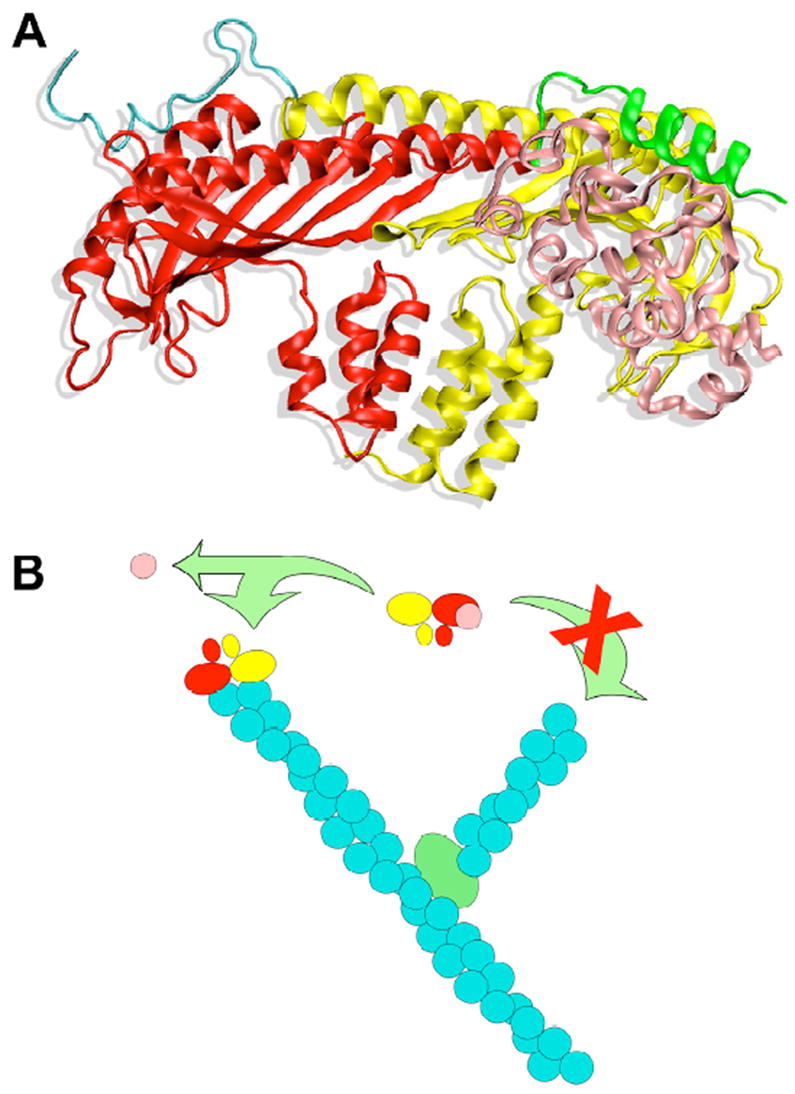
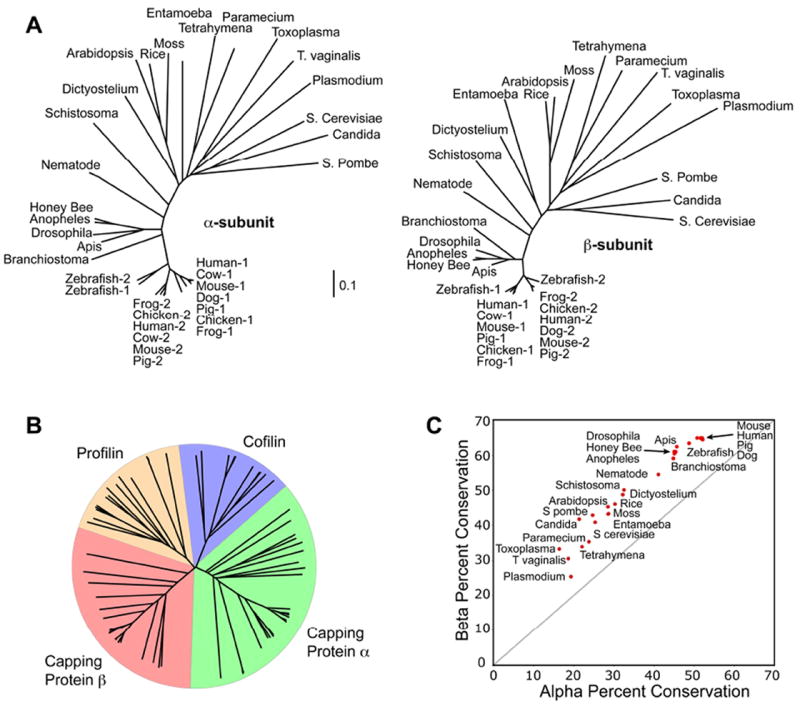
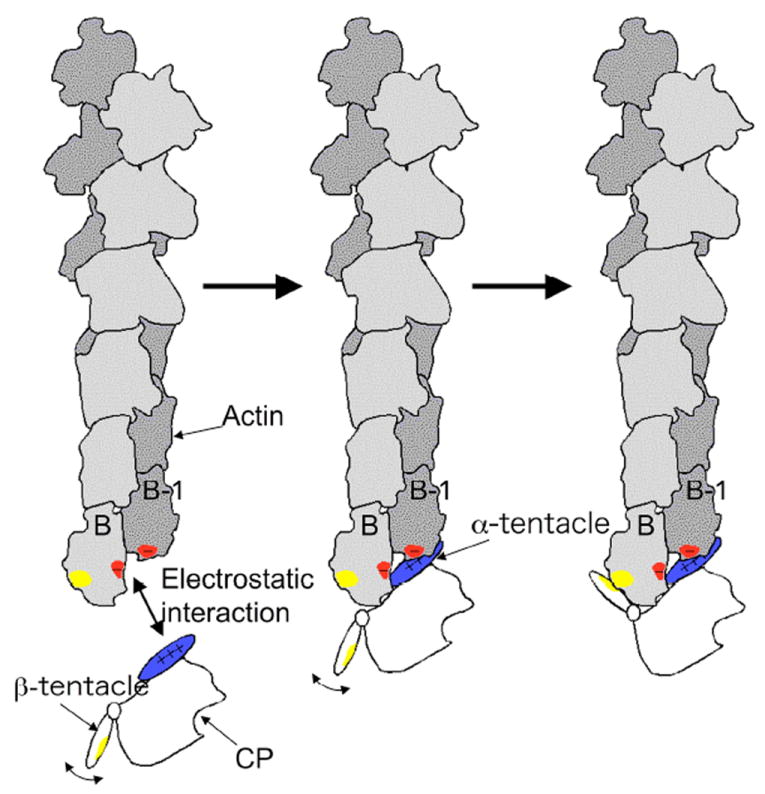
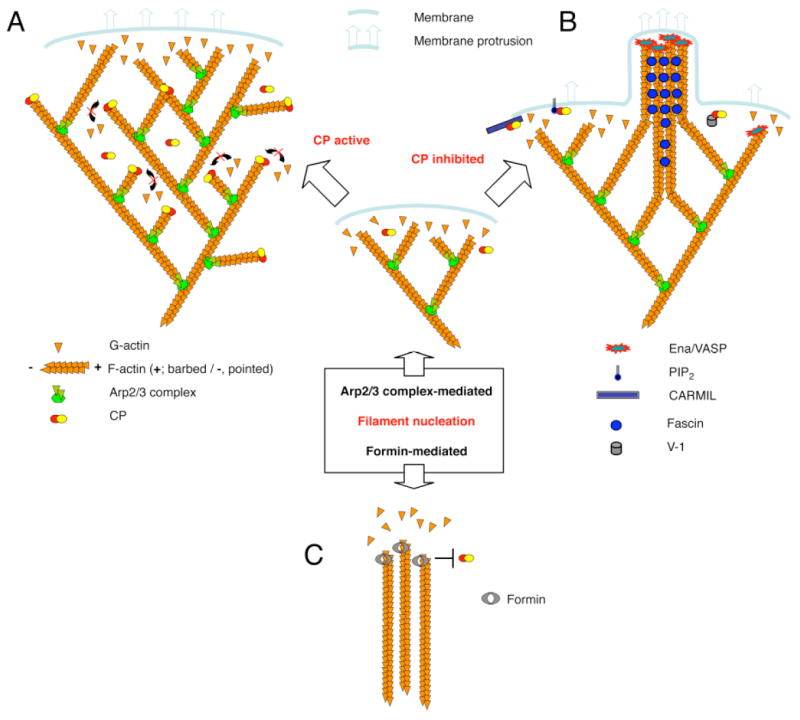
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
