

The p110beta isoform of phosphoinositide 3-kinase signals downstream of G protein-coupled receptors and is functionally redundant with p110gamma
- PMID: 18544649
- PMCID: PMC2448830
- DOI: 10.1073/pnas.0707761105
The p110beta isoform of phosphoinositide 3-kinase signals downstream of G protein-coupled receptors and is functionally redundant with p110gamma
Abstract
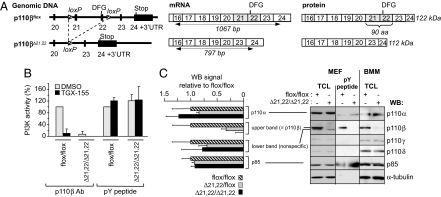
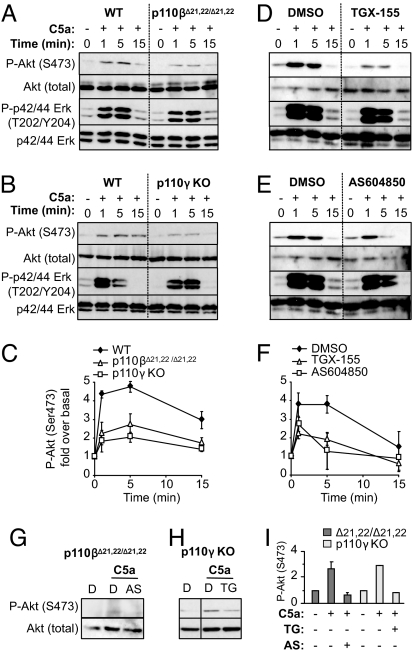
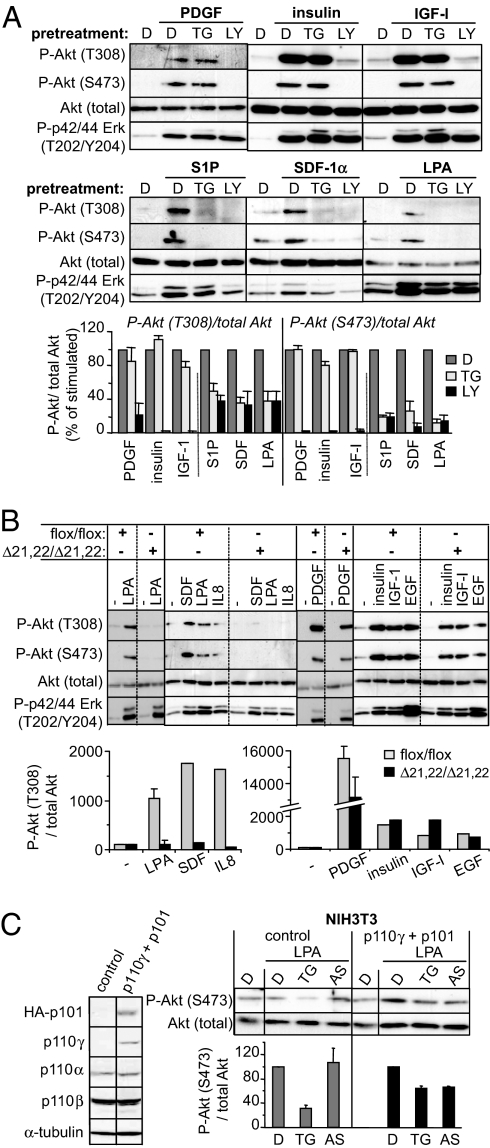
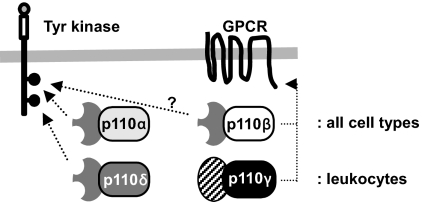
The p110 isoforms of phosphoinositide 3-kinase (PI3K) are acutely regulated by extracellular stimuli. The class IA PI3K catalytic subunits (p110alpha, p110beta, and p110delta) occur in complex with a Src homology 2 (SH2) domain-containing p85 regulatory subunit, which has been shown to link p110alpha and p110delta to Tyr kinase signaling pathways. The p84/p101 regulatory subunits of the p110gamma class IB PI3K lack SH2 domains and instead couple p110gamma to G protein-coupled receptors (GPCRs). Here, we show, using small-molecule inhibitors with selectivity for p110beta and cells derived from a p110beta-deficient mouse line, that p110beta is not a major effector of Tyr kinase signaling but couples to GPCRs. In macrophages, both p110beta and p110gamma contributed to Akt activation induced by the GPCR agonist complement 5a, but not by the Tyr kinase ligand colony-stimulating factor-1. In fibroblasts, which express p110beta but not p110gamma, p110beta mediated Akt activation by the GPCR ligands stromal cell-derived factor, sphingosine-1-phosphate, and lysophosphatidic acid but not by the Tyr kinase ligands PDGF, insulin, and insulin-like growth factor 1. Introduction of p110gamma in these cells reduced the contribution of p110beta to GPCR signaling. Taken together, these data show that p110beta and p110gamma can couple redundantly to the same GPCR agonists. p110beta, which shows a much broader tissue distribution than the leukocyte-restricted p110gamma, could thus provide a conduit for GPCR-linked PI3K signaling in the many cell types where p110gamma expression is low or absent.
Conflict of interest statement
Conflict of interest: B.V. is a consultant for PIramed (Slough, UK) and AstraZeneca.
Figures
References
-
- Vanhaesebroeck B, Leevers SJ, Panayotou G, Waterfield MD. Phosphoinositide 3-kinases: A conserved family of signal transducers. Trends Biochem Sci. 1997;22:267–272. - PubMed
-
- Bi L, Okabe I, Bernard DJ, Nussbaum RL. Early embryonic lethality in mice deficient in the p110β catalytic subunit of PI 3-kinase. Mamm Genome. 2002;13:169–172. - PubMed
-
- Jackson SP, et al. PI 3-kinase p110β: A new target for antithrombotic therapy. Nat Med. 2005;11:507–514. - PubMed
-
- Kurosu H, et al. Heterodimeric phosphoinositide 3-kinase consisting of p85 and p110β is synergistically activated by the β-γ subunits of G proteins and phosphotyrosyl peptide. J Biol Chem. 1997;272:24252–24256. - PubMed
-
- Maier U, Babich A, Nurnberg B. Roles of noncatalytic subunits in gbetagamma-induced activation of class I phosphoinositide 3-kinase isoforms β and γ. J Biol Chem. 1999;274:29311–29317. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- BB/C505659/2/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/C50989/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/C509890/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/B/0000M979/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/C505659/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
