

Inducible ablation of melanopsin-expressing retinal ganglion cells reveals their central role in non-image forming visual responses
- PMID: 18545654
- PMCID: PMC2396502
- DOI: 10.1371/journal.pone.0002451
Inducible ablation of melanopsin-expressing retinal ganglion cells reveals their central role in non-image forming visual responses
Erratum in
- PLoS ONE. 2008 Jun;4(6). doi: 10.1371/annotation/c02106ba-b00b-4416-9834-cf0f3ba49a37. Buch, Thorsten [added]; Waisman, Ari [added]
- PLoS ONE. 2008;3(7). doi: 10.1371/annotation/16f913dd-c33b-419f-9555-c788c80c189f
Abstract
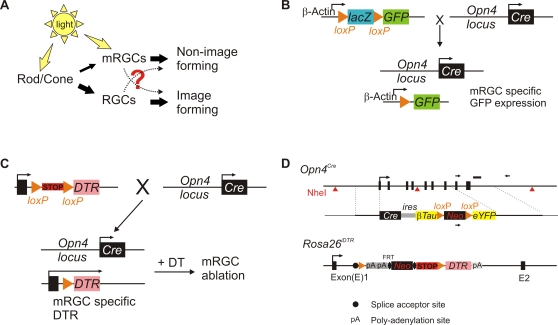
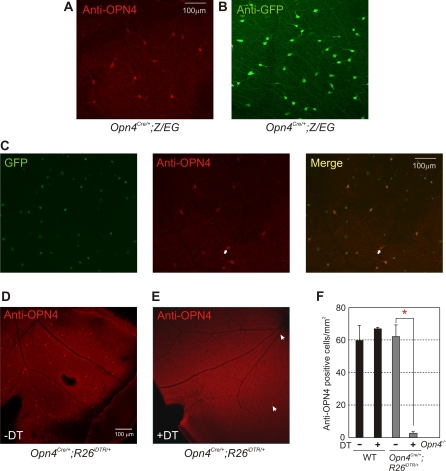
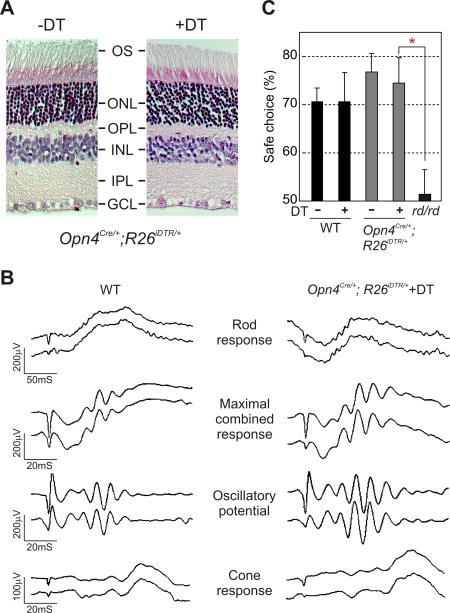
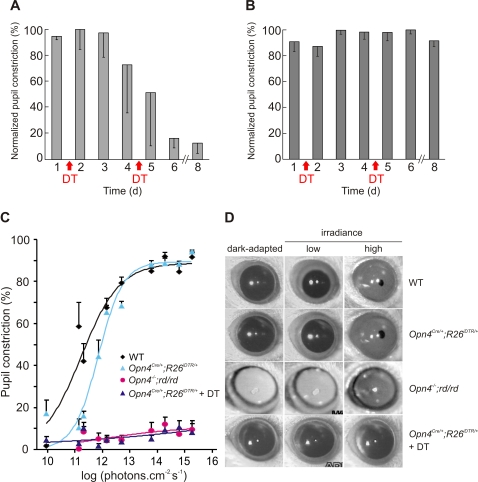
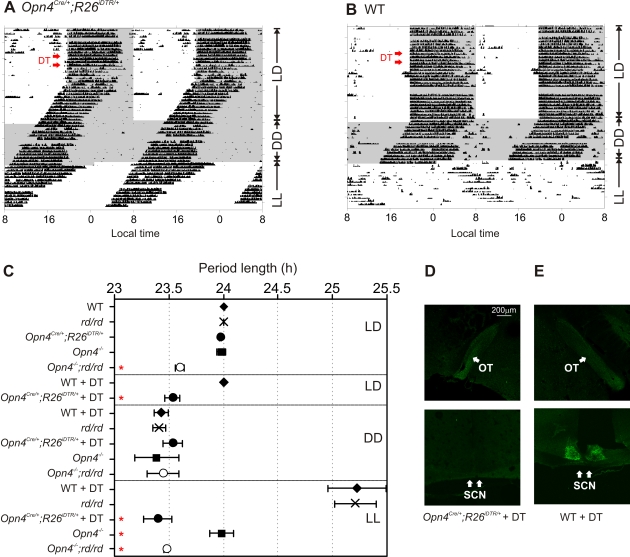
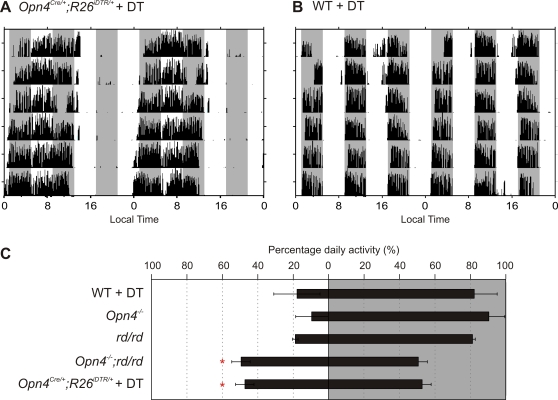
Rod/cone photoreceptors of the outer retina and the melanopsin-expressing retinal ganglion cells (mRGCs) of the inner retina mediate non-image forming visual responses including entrainment of the circadian clock to the ambient light, the pupillary light reflex (PLR), and light modulation of activity. Targeted deletion of the melanopsin gene attenuates these adaptive responses with no apparent change in the development and morphology of the mRGCs. Comprehensive identification of mRGCs and knowledge of their specific roles in image-forming and non-image forming photoresponses are currently lacking. We used a Cre-dependent GFP expression strategy in mice to genetically label the mRGCs. This revealed that only a subset of mRGCs express enough immunocytochemically detectable levels of melanopsin. We also used a Cre-inducible diphtheria toxin receptor (iDTR) expression approach to express the DTR in mRGCs. mRGCs develop normally, but can be acutely ablated upon diphtheria toxin administration. The mRGC-ablated mice exhibited normal outer retinal function. However, they completely lacked non-image forming visual responses such as circadian photoentrainment, light modulation of activity, and PLR. These results point to the mRGCs as the site of functional integration of the rod/cone and melanopsin phototransduction pathways and as the primary anatomical site for the divergence of image-forming and non-image forming photoresponses in mammals.
Conflict of interest statement
Figures
References
-
- Gooley JJ, Lu J, Chou TC, Scammell TE, Saper CB. Melanopsin in cells of origin of the retinohypothalamic tract. Nat Neurosci. 2001;4:1165. - PubMed
-
- Berson DM, Dunn FA, Takao M. Phototransduction by retinal ganglion cells that set the circadian clock. Science. 2002;295:1070–1073. - PubMed
-
- Belenky MA, Smeraski CA, Provencio I, Sollars PJ, Pickard GE. Melanopsin retinal ganglion cells receive bipolar and amacrine cell synapses. J Comp Neurol. 2003;460:380–393. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
