

Infection of mice with respiratory syncytial virus during neonatal life primes for enhanced antibody and T cell responses on secondary challenge
- PMID: 18549446
- PMCID: PMC2492908
- DOI: 10.1111/j.1365-2249.2008.03591.x
Infection of mice with respiratory syncytial virus during neonatal life primes for enhanced antibody and T cell responses on secondary challenge
Abstract
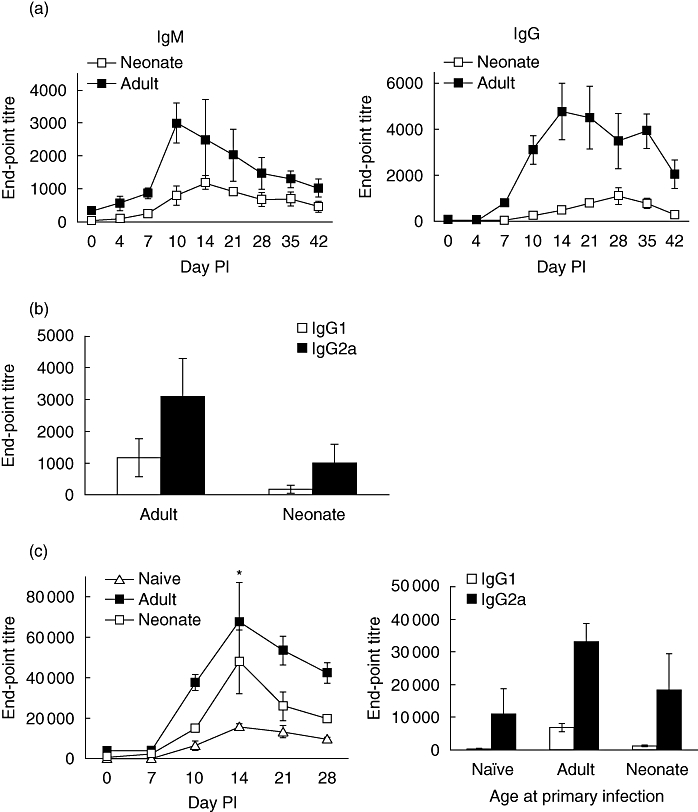
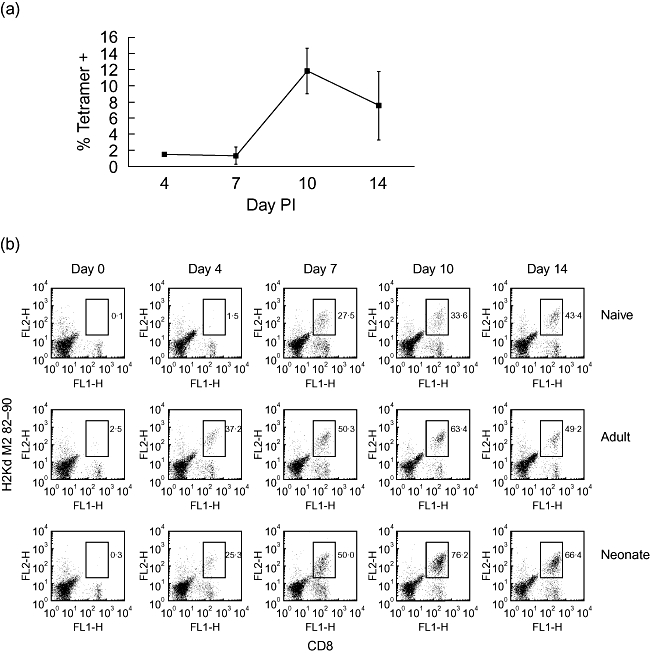
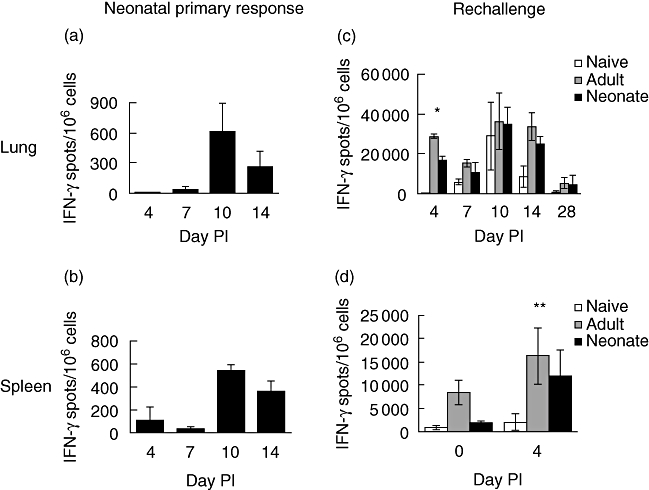
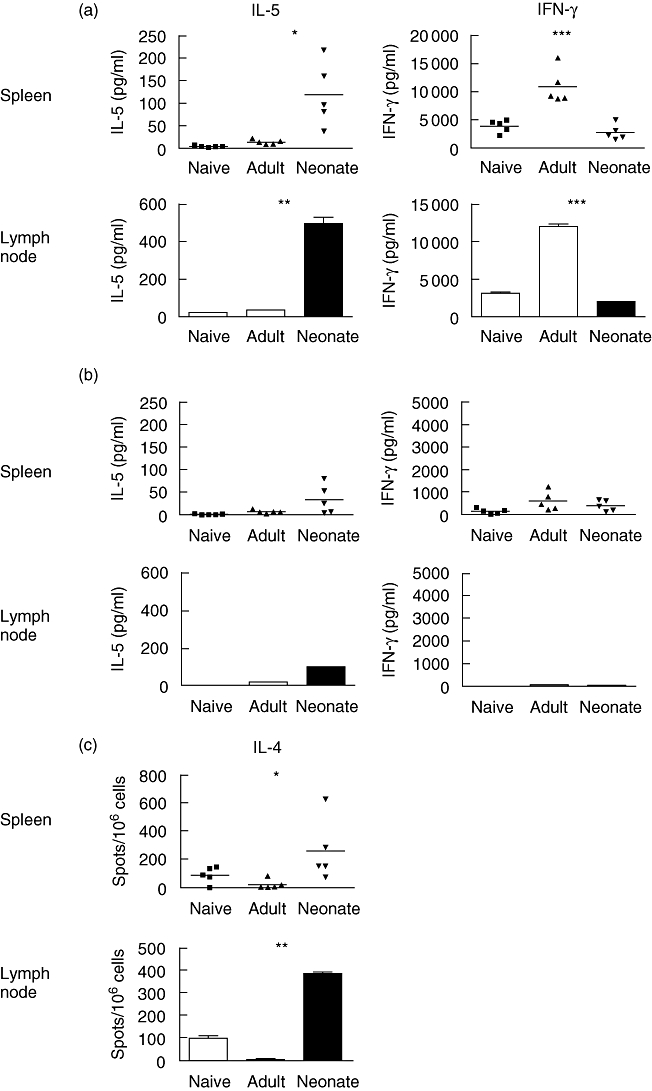
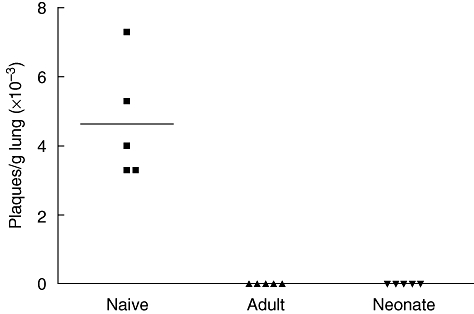
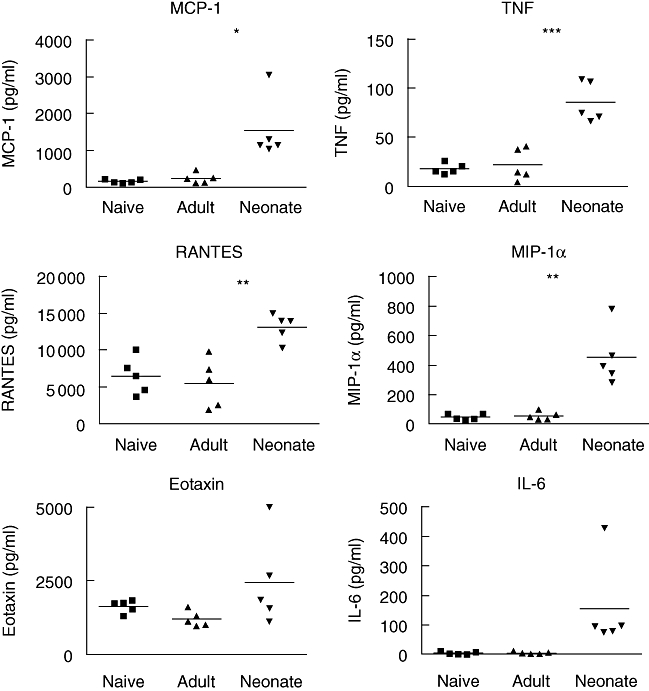
Primary neonatal immune responses to infection or vaccines are weak when compared with those of adults. In addition, memory responses of neonatally primed animals may be absent, weak or T helper type 2 (Th2)-biased. Respiratory syncytial virus (RSV) is an important pathogen of human infants and infection during the neonatal period has been linked to the development of asthma in later life. Here we report that acute intranasal infection of neonatal mice with RSV induces significant RSV-specific antibody and CD8 T cell responses. These responses were boosted after RSV rechallenge during adulthood, demonstrating the establishment of memory after neonatal priming. Primary infection during neonatal life was associated, following rechallenge, with limited viral replication in the lung. Recall responses of both spleen and lymph node cells from neonatally primed and adult-primed mice were associated with interferon-gamma secretion, indicative of a Th1-type response. However, interleukin (IL)-4 and IL-5 secretion were enhanced only in spleen and lymph node cells from neonatally primed mice. Rechallenge of neonatally primed mice was also associated with increased concentrations of chemokines monocyte chemoattractant protein-1, macrophage inflammatory protein-1alpha and regulated upon activation normal T cell expressed and secreted in the lung. These may play a role in the enhanced inflammatory cell recruitment and immunopathology induced following RSV reinfection. Our results demonstrate therefore that immunity to RSV can be established during neonatal life and, importantly, that the quality of the subsequent response is dependent upon the age of first infection.
Figures
References
-
- Billingham RE, Brent L, Medawar PB. Activity acquired tolerance of foreign cells. Nature. 1953;172:603–6. - PubMed
-
- Adkins B, Leclerc C, Marshall-Clarke S. Neonatal adaptive immunity comes of age. Nat Rev Immunol. 2004;4:553–64. - PubMed
-
- Marshall-Clarke S, Reen D, Tasker L, Hassan J. Neonatal immunity: how well has it grown up? Immunol Today. 2000;21:35–41. - PubMed
-
- Siegrist CA. Neonatal and early life vaccinology. Vaccine. 2001;19:3331–46. - PubMed
-
- Garcia AM, Fadel SA, Cao S, Sarzotti M. T cell immunity in neonates. Immunol Res. 2000;22:177–90. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
