

doi: 10.1128/JVI.02575-07.
Epub 2008 Jun 11.
Liver-specific microRNA miR-122 enhances the replication of hepatitis C virus in nonhepatic cells
Affiliations
- PMID: 18550664
- PMCID: PMC2519557
- DOI: 10.1128/JVI.02575-07
Item in Clipboard
Liver-specific microRNA miR-122 enhances the replication of hepatitis C virus in nonhepatic cells
J Virol.
2008 Aug.
Abstract
The liver-specific microRNA miR-122 has been shown to be required for the replication of hepatitis C virus (HCV) in the hepatoma cell line Huh7. The aim of this study was to test if HCV replication can be modulated by exogenously expressed miR-122 in human embryonic kidney epithelial cells (HEK-293). Our results demonstrate that miR-122 enhances the colony formation efficiency of the HCV replicon and increases the steady-state level of HCV RNA in HEK-293 cells. Therefore, we conclude that although miR-122 is not absolutely required, it greatly enhances HCV replication in nonhepatic cells.
Figures
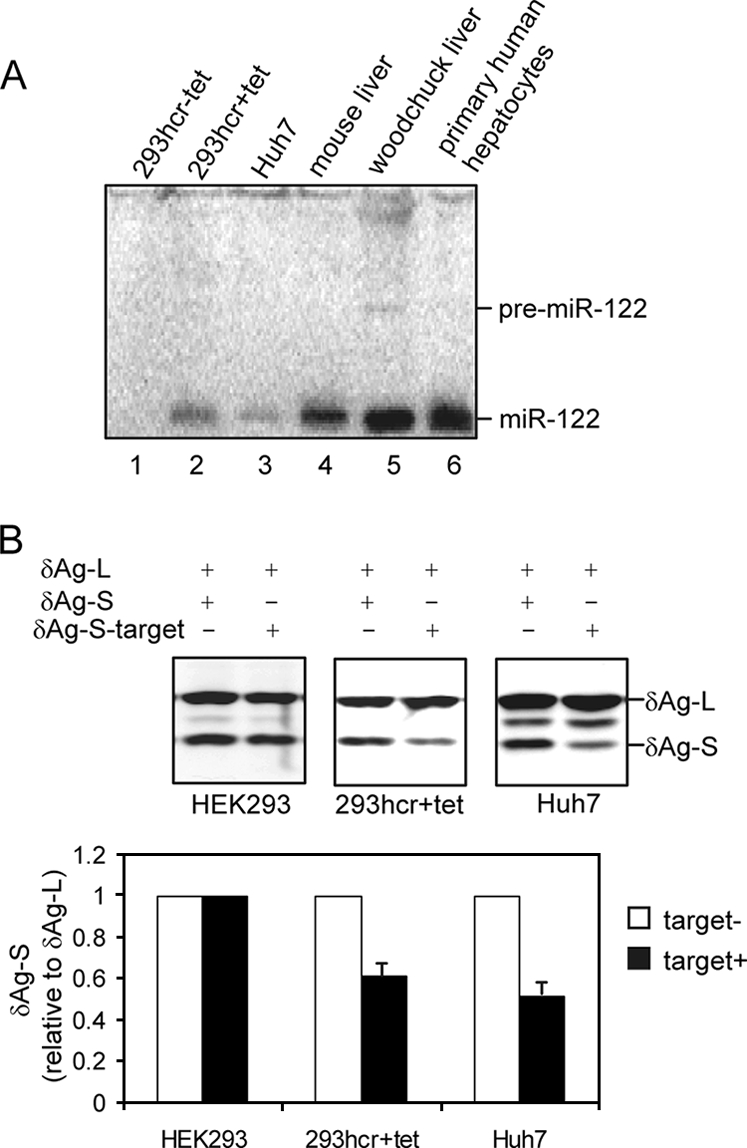
Establishment and characterization of 293 cells inducibly expressing miR-122. FLP-IN T Rex (Invitrogen) cells were cotransfected with plasmid pcDNA5/RFT/hcr that contains 160 bp of the woodchuck genomic hcr sequence encoding miR-122 and pOG44 to establish a stable cell line (293hcr) in which the transcription of miR-122 containing sequence is under the control of a tetracycline-inducible promoter. (A) 293hcr cells were cultured in the absence (lane 1) or presence (lane 2) of 1 μg/ml tetracycline for 3 days. Total RNA was extracted, and 10 μg RNA was used to detect miR-122 by Northern blot hybridization with a [γ-32P]ATP-labeled oligonucleotide that is complementary to miR-122. Ten-microgram portions of total RNA extracted from Huh7 cells (lane 3), mouse liver (lane 4), woodchuck liver (lane 5), and primary human hepatocytes (lane 6) served as controls. The positions of the ∼70-nt miR-122 precursor (pre-miR-122) and the ∼22-nt miR-122 are indicated. (B) The function of miR-122 processed from integrated hcr transcript was validated by a reporter assay described previously (10). 293 cells and 293hcr cells cultured in the presence of 1 μg/ml tetracycline and Huh7 cells were cotransfected with an internal control plasmid and either one of the two reporter plasmids. The internal control and reporter constructs expressed δAg-L and δAg-S, respectively. As indicated, in one reporter plasmid, a 22-nt miR-122 target sequence was inserted in the 3′ UTR (δAg-S-target). Cells were harvested 3 days after transfection, and total cellular protein was analyzed by Western blotting using a rabbit polyclonal antibody recognizing both forms of the δAg. The bound antibody was visualized by incubation with an infrared-dye-labeled goat anti-rabbit antibody (LI-COR) and quantified with an Odyssey apparatus (LI-COR).
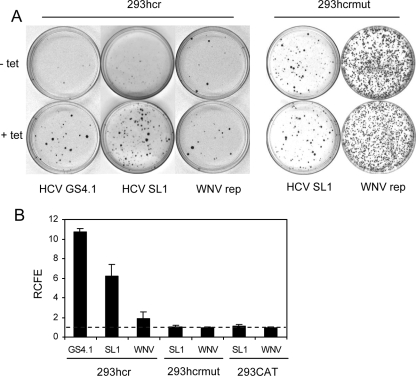
Cell colony formation efficiency of the HCV replicon was enhanced by exogenous expression of miR-122 in 293 cells. 293hcr, 293hcrmut, or 293CAT cells were cultured in the absence or presence of tetracycline for 2 days and then electroporated with total RNA extracted from HCV replicon-containing Huh7 (GS4.1) and HeLa (SL1) cells or WNV replicon-containing 293 cells (WNVrep), respectively. Transfected cells were then cultured in the absence or presence of tetracycline and selected with 500 μg/ml of G-418 for 2 to 3 weeks (see the text for details). (A) Cell foci were stained with crystal violet, and representative photographs of 293hcr and 293hcrmut cells are shown. (B) The cell foci in three plates cultured in either the absence or presence of tetracycline were counted. RCFE was expressed and plotted as the ratio of the number of foci obtained from cells selected in the presence of tetracycline to that from cells cultured in the absence of the antibiotic.
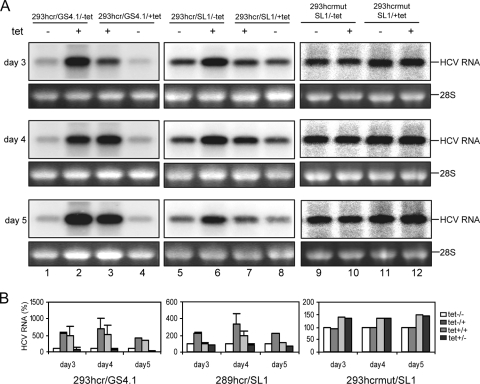
Steady-state replication of HCV replicons was enhanced by exogenous expression of miR-122 in 293 cells. (A) 293hcr- and 293hcrmut-derived HCV replicon-containing cell lines established in the absence (293hcr/GS4.1/-tet, 293hcr/SL1/-tet and 293hcrmut/SL1/-tet) or presence (293hcr/GS4.1/+tet, 293hcr/SL1/+tet and 293hcrmut/SL1/+tet) of tetracycline were seeded in 12-well plates at a density of 5 × 104 cells per well. Cells were either cultured under the original conditions or switched to medium with or without tetracycline. Cells were then harvested at the indicated time points after seeding, and total cellular RNA was extracted. Ten micrograms of total RNAs was analyzed by Northern blot analysis with an [α-32P]UTP-labeled riboprobe that is complementary to the plus strand of the HCV NS3 coding region. 28S rRNA served as the loading control. (B) Levels of viral RNA were quantified with a Bio-Rad Bioimager, and the mean values and standard deviations from three independent experiments were plotted.
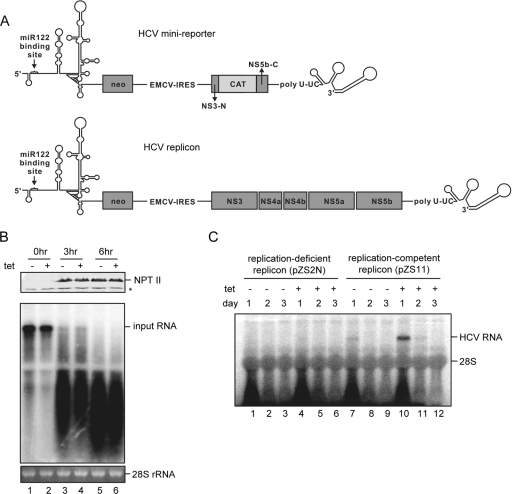
Exogenous expression of miR-122 in 293 cells does not affect HCV RNA stability and translation activity but dramatically enhances viral RNA replication. (A) Schematic representation of HCV minireporter (top) and dicistronic HCV replicon (49). The cis element in NS5B that forms a kissing loop with the 3′ UTR sequence is retained in the minireporter. (B) HCV minireporter RNA with significant deletion of the nonstructural protein coding region was electroporated into 293hcr cells without or with 2 days of prior tetracycline induction, by following the protocol described previously (19). Transfected cells were seeded in 60-mm dishes and continued to be cultured in the absence or presence of tetracycline for the indicated period. HCV minireporter RNA was analyzed by Northern blot hybridization with a riboprobe that is complementary to the plus strand of the NPT II coding region. 28S rRNA served as the loading control (bottom). NPT II protein in cell lysates was analyzed by Western blotting (top) using a rabbit polyclonal antibody (Upstate). The bound antibody was visualized by incubation with an infrared-dye-labeled goat anti-rabbit antibody (LI-COR). The asterisk indicates a cross-reaction band. (C) An in vitro-transcribed replication-competent HCV replicon (pZS11) or a replication-deficient replicon with a 2-nt in-frame deletion (pZS2N) was electroporated into 293hcr cells without or with 2 days of prior tetracycline induction. Transfected cells were harvested at 1, 2, and 3 day posttransfection. Ten micrograms of total cellular RNA were resolved in 1% agarose gel containing 2.2 M formaldehyde and transferred onto Nylon membrane. The membrane was probed with a [α-32P]UTP-labeled riboprobe that is complementary to the plus strand of the HCV NS3 coding region.
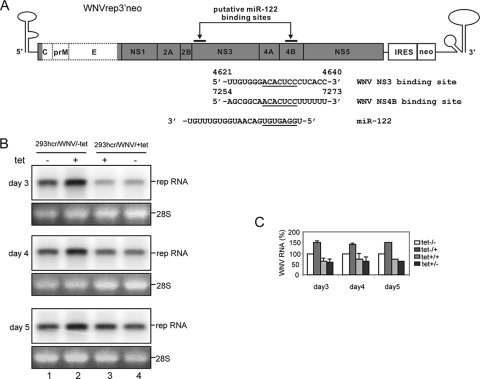
Effect of miR-122 on the replication of the WNV subgenomic replicon. (A) Schematic representation of the WNV subgenomic replicon (WNVrep3′neo) (42) and the putative miR-122 target sites and sequences, with seed and seed-match sequences underlined (numbering is according to a published WNV sequence, GenBank accession number AF404756). (B) 293hcr-derived WNV replicon-containing cell lines established in the absence (293hcr/WNV/-tet) and presence (293hcr/WNV/+tet) of tetracycline were seeded in 12-well plates at a density of 5 × 104 cells per well. Cells were either cultured under the original conditions or switched to medium with or without tetracycline. Cells were then harvested at the indicated time points after seeding, and total cellular RNA was extracted. Ten micrograms of total RNA was analyzed by Northern blot analysis with an [α-32P]UTP-labeled riboprobe that is complementary to the plus strand of the NPT II coding region. 28S rRNA served as the loading control. (C) Levels of viral RNA were quantified with a Bio-Rad Bioimager, and the mean values and standard deviations from three independent experiments were plotted.
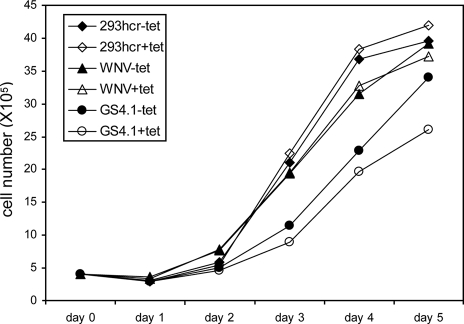
Effects of miR-122 expression and viral replication on cell growth. The indicated cell lines (293hcr, WNV, and HCV GS4.1) were seeded in six-well plates at a density of 5 × 105 cells per well in medium with or without 1 μg/ml tetracycline. Three wells of cells from each of the cell lines cultured under either condition were trypsinized at 1, 2, 3, 4, and 5 days after seeding. Average cell numbers from the three wells were plotted.
References
-
- Akira, S., and K. Takeda. 2004. Toll-like receptor signalling. Nat. Rev. Immunol. 4499-511. - PubMed
-
- Ambros, V. 2004. The functions of animal microRNAs. Nature 431350-355. - PubMed
-
- Bartel, D. P. 2004. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 116281-297. - PubMed
-
- Berkhout, B., and K. T. Jeang. 2007. RISCy business: microRNAs, pathogenesis, and viruses. J. Biol. Chem. 28226641-26645. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
