

CpG dinucleotides and the mutation rate of non-CpG DNA
- PMID: 18550801
- PMCID: PMC2527700
- DOI: 10.1101/gr.076455.108
CpG dinucleotides and the mutation rate of non-CpG DNA
Abstract
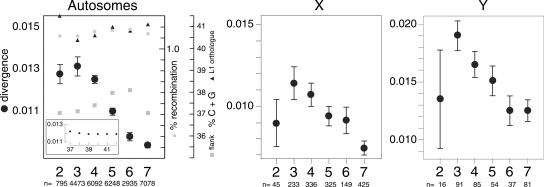
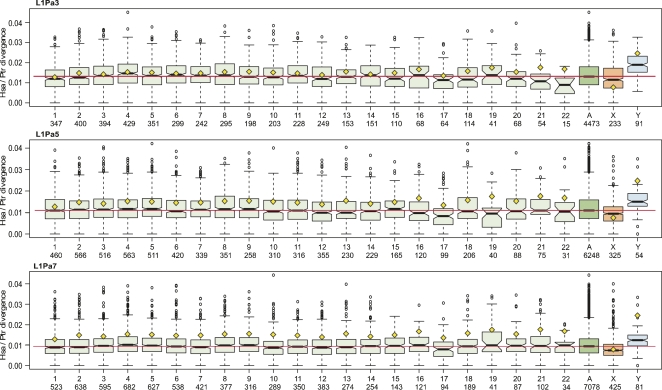
The neutral mutation rate is equal to the base substitution rate when the latter is not affected by natural selection. Differences between these rates may reveal that factors such as natural selection, linkage, or a mutator locus are affecting a given sequence. We examined the neutral base substitution rate by measuring the sequence divergence of approximately 30,000 pairs of inactive orthologous L1 retrotransposon sequences interspersed throughout the human and chimpanzee genomes. In contrast to other studies, we related ortholog divergence to the time (age) that the L1 sequences resided in the genome prior to the chimpanzee and human speciation. As expected, the younger orthologs contained more hypermutable CpGs than the older ones because of their conversion to TpGs (and CpAs). Consequently, the younger orthologs accumulated more CpG mutations than the older ones during the approximately 5 million years since the human and chimpanzee lineages separated. But during this same time, the younger orthologs also accumulated more non-CpG mutations than the older ones. In fact, non-CpG and CpG mutations showed an almost perfect (R2 = 0.98) correlation for approximately 97% of the ortholog pairs. The correlation is independent of G + C content, recombination rate, and chromosomal location. Therefore, it likely reflects an intrinsic effect of CpGs, or mutations thereof, on non-CpG DNA rather than the joint manifestation of the chromosomal environment. The CpG effect is not uniform for all regions of non-CpG DNA. Therefore, the mutation rate of non-CpG DNA is contingent to varying extents on local CpG content. Aside from their implications for mutational mechanisms, these results indicate that a precise determination of a uniform genome-wide neutral mutation rate may not be attainable.
Figures




References
-
- Adey N.B., Tollefsbol T.O., Sparks A.B., Edgell M.H., Hutchison C.A.I., Tollefsbol T.O., Sparks A.B., Edgell M.H., Hutchison C.A.I., Sparks A.B., Edgell M.H., Hutchison C.A.I., Edgell M.H., Hutchison C.A.I., Hutchison C.A.I. Molecular resurrection of an extinct ancestral promoter for mouse L1. Proc. Natl. Acad. Sci. 1994;91:1569–1573. - PMC - PubMed
-
- Asthana S., Schmidt S., Sunyaev S., Schmidt S., Sunyaev S., Sunyaev S. A limited role for balancing selection. Trends Genet. 2005;21:30–32. - PubMed
-
- Bird A. DNA methylation patterns and epigenetic memory. Genes & Dev. 2002;16:6–21. - PubMed
-
- Bohossian H.B., Skaletsky H., Page D.C., Skaletsky H., Page D.C., Page D.C. Unexpected similar rates of nucleotide substitution found in male and female hominids. Nature. 2000;406:622–625. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials