

Mitochondrial morphogenesis, dendrite development, and synapse formation in cerebellum require both Bcl-w and the glutamate receptor delta2
- PMID: 18551174
- PMCID: PMC2405952
- DOI: 10.1371/journal.pgen.1000097
Mitochondrial morphogenesis, dendrite development, and synapse formation in cerebellum require both Bcl-w and the glutamate receptor delta2
Abstract
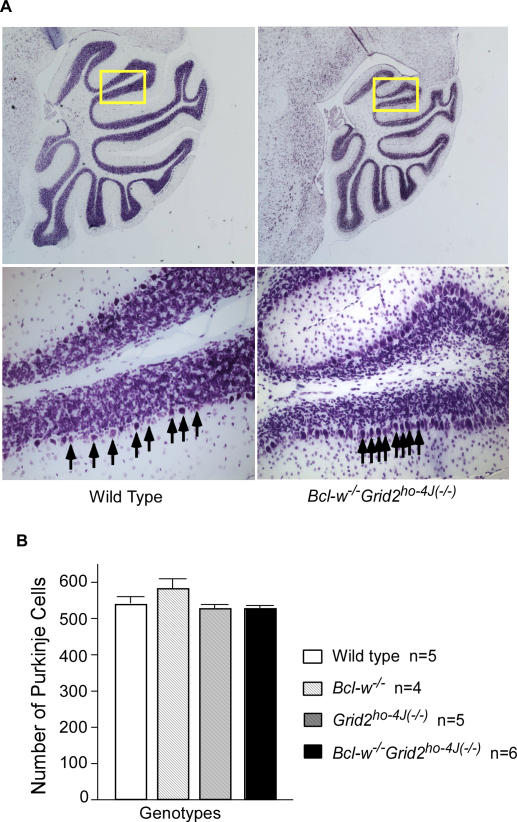
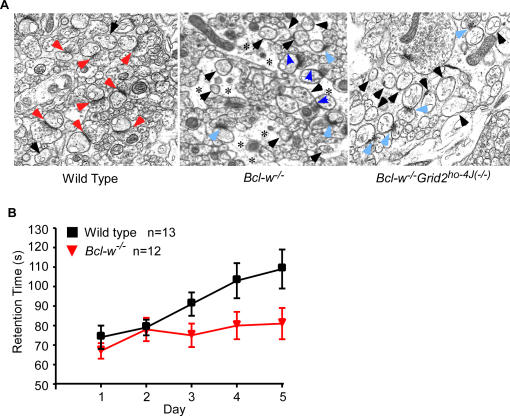
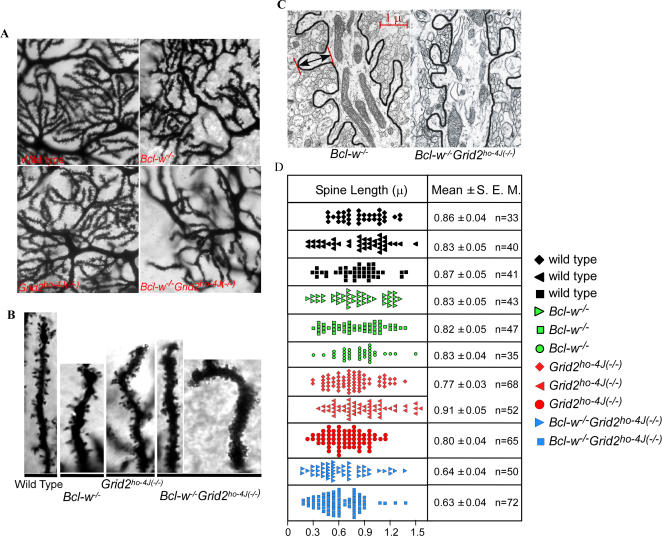
Bcl-w belongs to the prosurvival group of the Bcl-2 family, while the glutamate receptor delta2 (Grid2) is an excitatory receptor that is specifically expressed in Purkinje cells, and required for Purkinje cell synapse formation. A recently published result as well as our own findings have shown that Bcl-w can physically interact with an autophagy protein, Beclin1, which in turn has been shown previously to form a protein complex with the intracellular domain of Grid2 and an adaptor protein, nPIST. This suggests that Bcl-w and Grid2 might interact genetically to regulate mitochondria, autophagy, and neuronal function. In this study, we investigated this genetic interaction of Bcl-w and Grid2 through analysis of single and double mutant mice of these two proteins using a combination of histological and behavior tests. It was found that Bcl-w does not control the cell number in mouse brain, but promotes what is likely to be the mitochondrial fission in Purkinje cell dendrites, and is required for synapse formation and motor learning in cerebellum, and that Grid2 has similar phenotypes. Mice carrying the double mutations of these two genes had synergistic effects including extremely long mitochondria in Purkinje cell dendrites, and strongly aberrant Purkinje cell dendrites, spines, and synapses, and severely ataxic behavior. Bcl-w and Grid2 mutations were not found to influence the basal autophagy that is required for Purkinje cell survival, thus resulting in these phenotypes. Our results demonstrate that Bcl-w and Grid2 are two critical proteins acting in distinct pathways to regulate mitochondrial morphogenesis and control Purkinje cell dendrite development and synapse formation. We propose that the mitochondrial fission occurring during neuronal growth might be critically important for dendrite development and synapse formation, and that it can be regulated coordinately by multiple pathways including Bcl-2 and glutamate receptor family members.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures


References
-
- Polyakov VY, Soukhomlinova MY, Fais D. Fusion, fragmentation, and fission of mitochondria. Biochemistry (Mosc) 2003;68:838–849. - PubMed
-
- Okamoto K, Shaw JM. Mitochondrial morphology and dynamics in yeast and multicellular eukaryotes. Annu Rev Genet. 2005;39:503–536. - PubMed
-
- Chan DC. Mitochondrial Fusion and Fission in Mammals. Annu Rev Cell Dev Biol. 2006;22:79–99. - PubMed
-
- Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P. Molecular Biology of the Cell (4th edition) GS Garland Science Taylor & Francis group; 2002. pp. 808–821.
-
- Youle RJ, Karbowski M. Mitochondrial fission in apoptosis. Nat Rev Mol Cell Biol. 2005;6:657–663. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
