

Dishevelled controls apical docking and planar polarization of basal bodies in ciliated epithelial cells
- PMID: 18552847
- PMCID: PMC2771675
- DOI: 10.1038/ng.104
Dishevelled controls apical docking and planar polarization of basal bodies in ciliated epithelial cells
Abstract
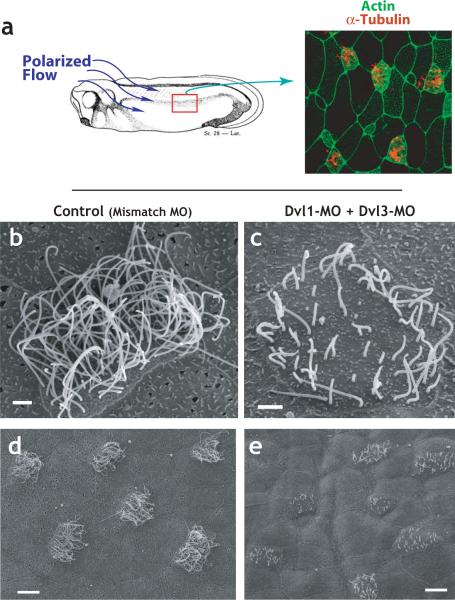
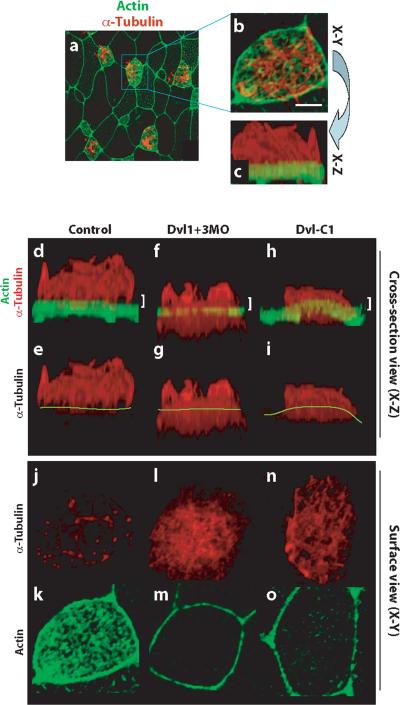
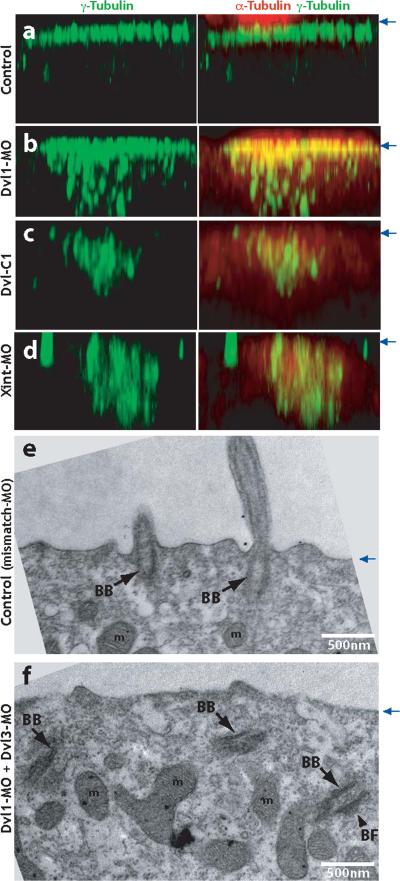
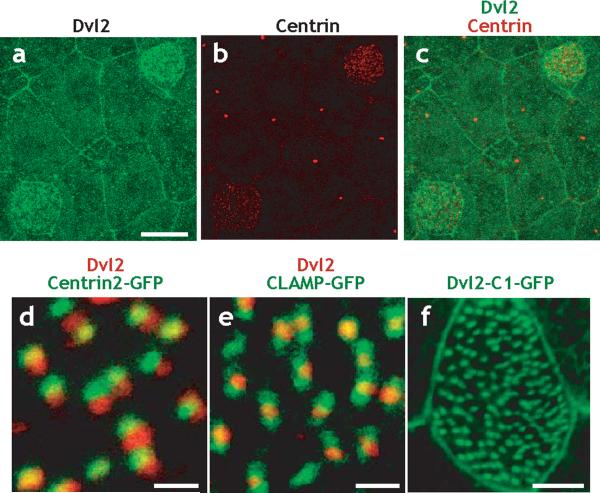
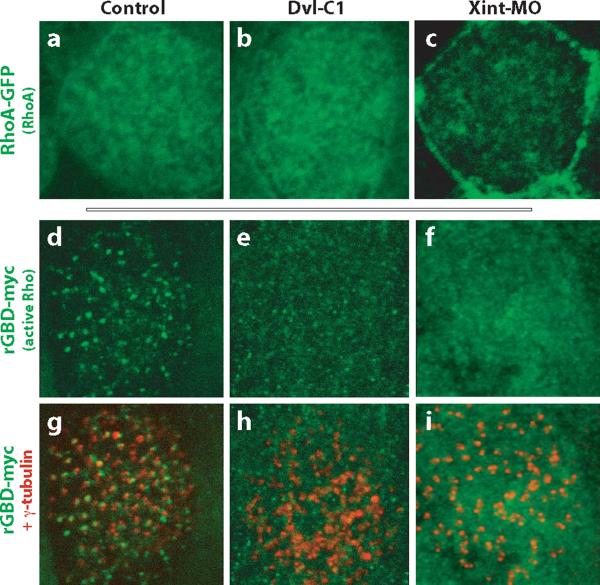
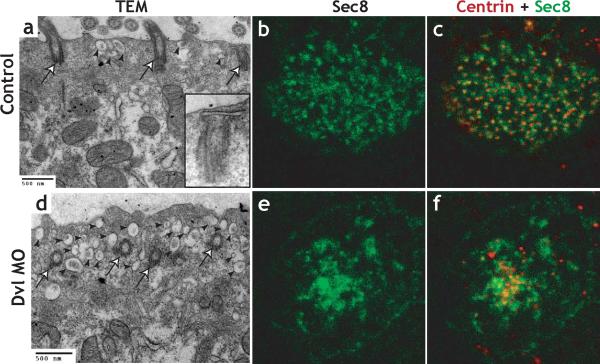
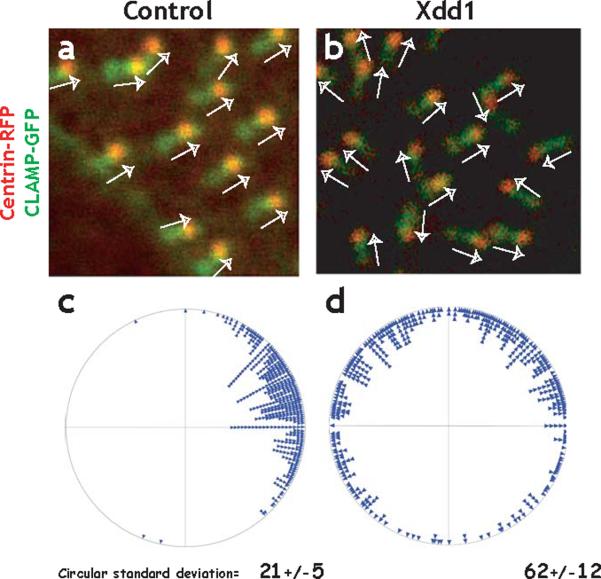
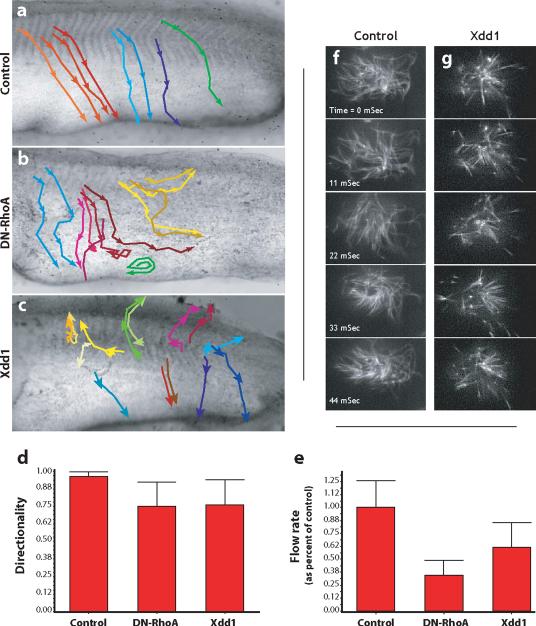
The planar cell polarity (PCP) signaling system governs many aspects of polarized cell behavior. Here, we use an in vivo model of vertebrate mucociliary epithelial development to show that Dishevelled (Dvl) is essential for the apical positioning of basal bodies. We find that Dvl and Inturned mediate the activation of the Rho GTPase specifically at basal bodies, and that these three proteins together mediate the docking of basal bodies to the apical plasma membrane. Moreover, we find that this docking involves a Dvl-dependent association of basal bodies with membrane-bound vesicles and the vesicle-trafficking protein, Sec8. Once docked, basal bodies again require Dvl and Rho for the planar polarization that underlies directional beating of cilia. These results demonstrate previously undescribed functions for PCP signaling components and suggest that a common signaling apparatus governs both apical docking and planar polarization of basal bodies.
Figures
References
-
- Chilvers MA, O'Callaghan C. Local mucociliary defence mechanisms. Paediatr Respir Rev. 2000;1:27–34. - PubMed
-
- Steinman RM. An electron microscopic study of ciliogenesis in developing epidermis and trachea in the embryo of Xenopus laevis. Am J Anat. 1968;122:19–55. - PubMed
-
- Raji AR, Naserpour M. Light and Electron Microscopic Studies of the Trachea in the One-Humped Camel (Camelus dromedarius) Anat Histol Embryol. 2007;36:10–3. - PubMed
-
- Dawe HR, Farr H, Gull K. Centriole/basal body morphogenesis and migration during ciliogenesis in animal cells. J Cell Sci. 2007;120:7–15. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
