

EsaC substrate for the ESAT-6 secretion pathway and its role in persistent infections of Staphylococcus aureus
- PMID: 18554323
- PMCID: PMC2597432
- DOI: 10.1111/j.1365-2958.2008.06324.x
EsaC substrate for the ESAT-6 secretion pathway and its role in persistent infections of Staphylococcus aureus
Abstract
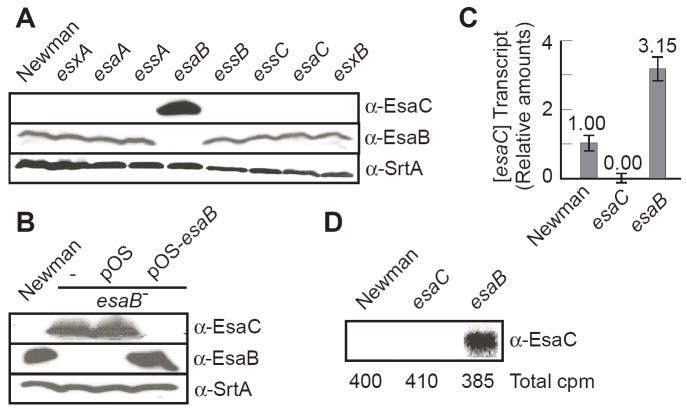
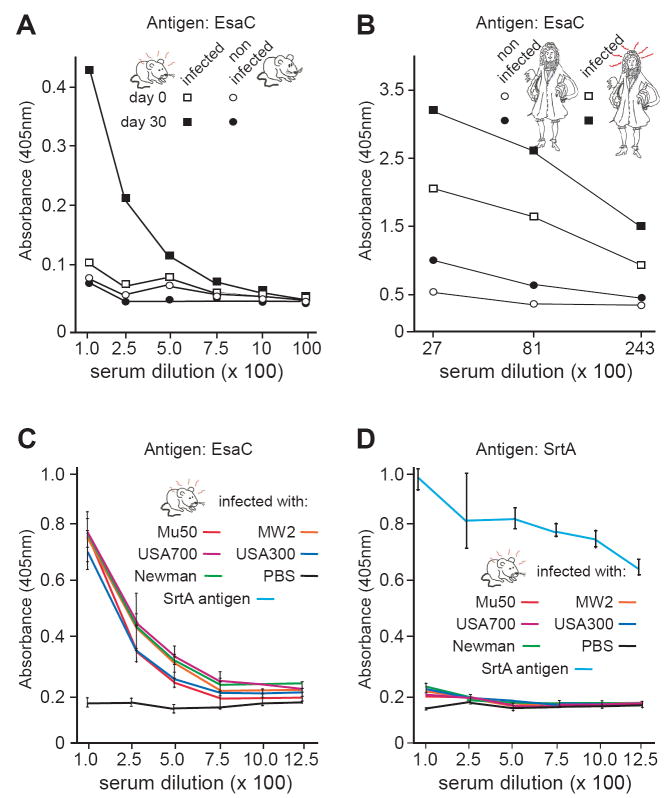
Staphylococcus aureus encodes the specialized secretion system Ess (ESAT-6 secretion system). The ess locus is a cluster of eight genes (esxAB, essABC, esaABC) of which esxA and esxB display homology to secreted ESAT-6 proteins of Mycobacterium tuberculosis. EsxA and EsxB require EssA, EssB and EssC for transport across the staphylococcal envelope. Herein, we examine the role of EsaB and EsaC and show that EsaB is a negative regulator of EsaC. Further, EsaC production is repressed when staphylococci are grown in broth and increased when staphylococci replicate in serum or infected hosts. EsaB is constitutively produced and remains in the cytoplasm whereas EsaC is secreted. This secretion requires an intact Ess pathway. Mutants lacking esaB or esaC display only a small defect in acute infection, but remarkably are unable to promote persistent abscesses during animal infection. Together, the data suggest a model whereby EsaB controls the production of effector molecules that are important for host pathogen interaction. One such effector, EsaC, is a secretion substrate of the Ess pathway and implements its pathogenic function during infection.
Figures





References
-
- Abdallah AM, Verboom T, Hannes F, Safi M, Strong M, Eisenberg D, Musters RJ, Vandenbroucke-Grauls CM, Appelmelk BJ, Luirink J, Bitter W. A specific secretion system mediates PPE41 transport in pathogenic mycobacteria. Mol Microbiol. 2006;62:667–679. - PubMed
-
- Abdallah AM, Gey van Pittius NC, Champion PA, Cox J, Luirink J, Vandenbroucke-Grauls CM, Appelmelk BJ, Bitter W. Type VII secretion--mycobacteria show the way. Nat Rev Microbiol. 2007;5:883–891. - PubMed
-
- Andersen P, Andersen AB, Sorensen AL, Nagai S. Recall of long-lived immunity to Mycobacterium tuberculosis infection in mice. J Immunol. 1995;154:3359–3372. - PubMed
-
- Archer GL. Staphylococcus aureus: a well-armed pathogen. Clin Infect Dis. 1998;26:1179–1181. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
