

The amyotrophic lateral sclerosis 8 protein VAPB is cleaved, secreted, and acts as a ligand for Eph receptors
- PMID: 18555774
- PMCID: PMC2494862
- DOI: 10.1016/j.cell.2008.04.039
The amyotrophic lateral sclerosis 8 protein VAPB is cleaved, secreted, and acts as a ligand for Eph receptors
Abstract
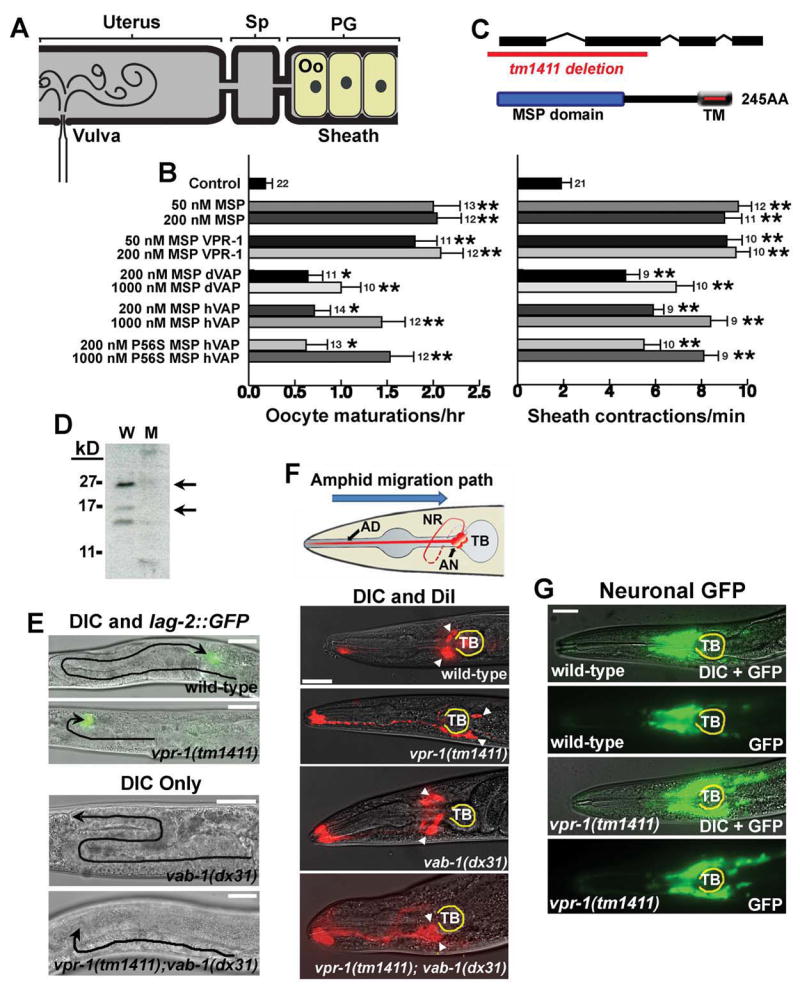
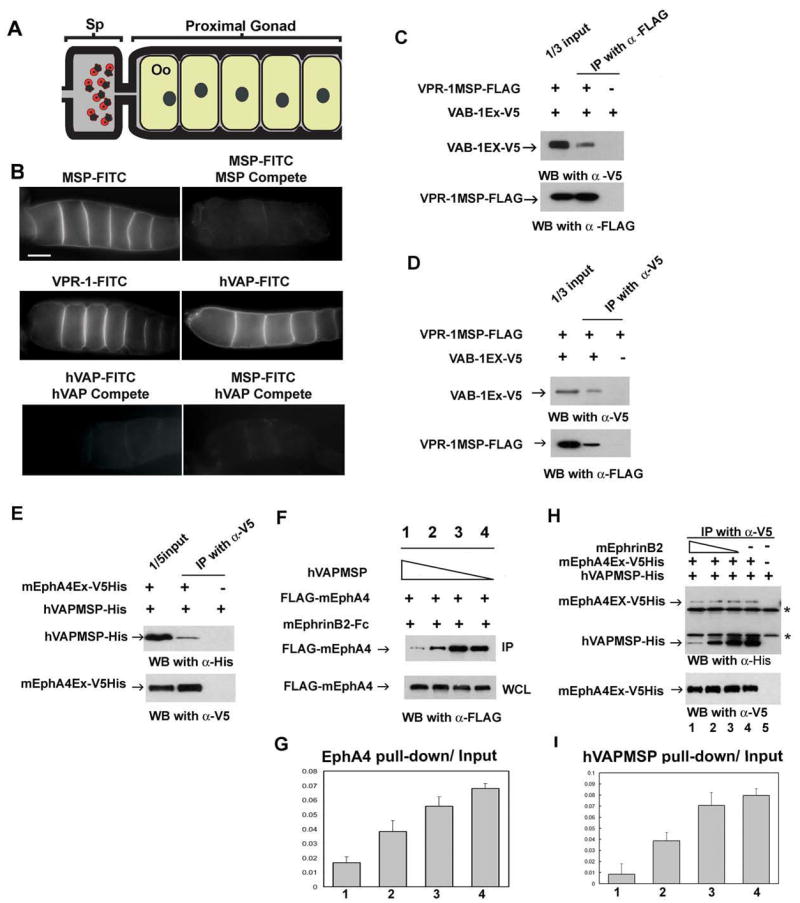
VAP proteins (human VAPB/ALS8, Drosophila VAP33, and C. elegans VPR-1) are homologous proteins with an amino-terminal major sperm protein (MSP) domain and a transmembrane domain. The MSP domain is named for its similarity to the C. elegans MSP protein, a sperm-derived hormone that binds to the Eph receptor and induces oocyte maturation. A point mutation (P56S) in the MSP domain of human VAPB is associated with Amyotrophic lateral sclerosis (ALS), but the mechanisms underlying the pathogenesis are poorly understood. Here we show that the MSP domains of VAP proteins are cleaved and secreted ligands for Eph receptors. The P58S mutation in VAP33 leads to a failure to secrete the MSP domain as well as ubiquitination, accumulation of inclusions in the endoplasmic reticulum, and an unfolded protein response. We propose that VAP MSP domains are secreted and act as diffusible hormones for Eph receptors. This work provides insight into mechanisms that may impact the pathogenesis of ALS.
Figures

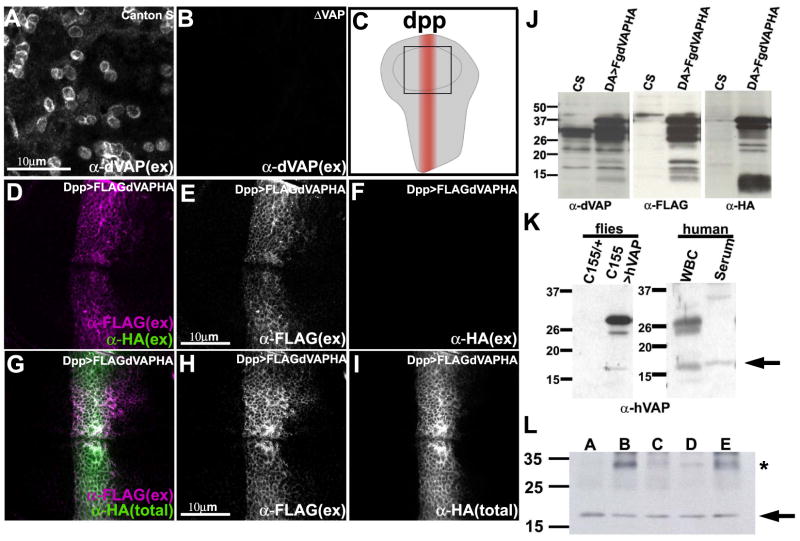
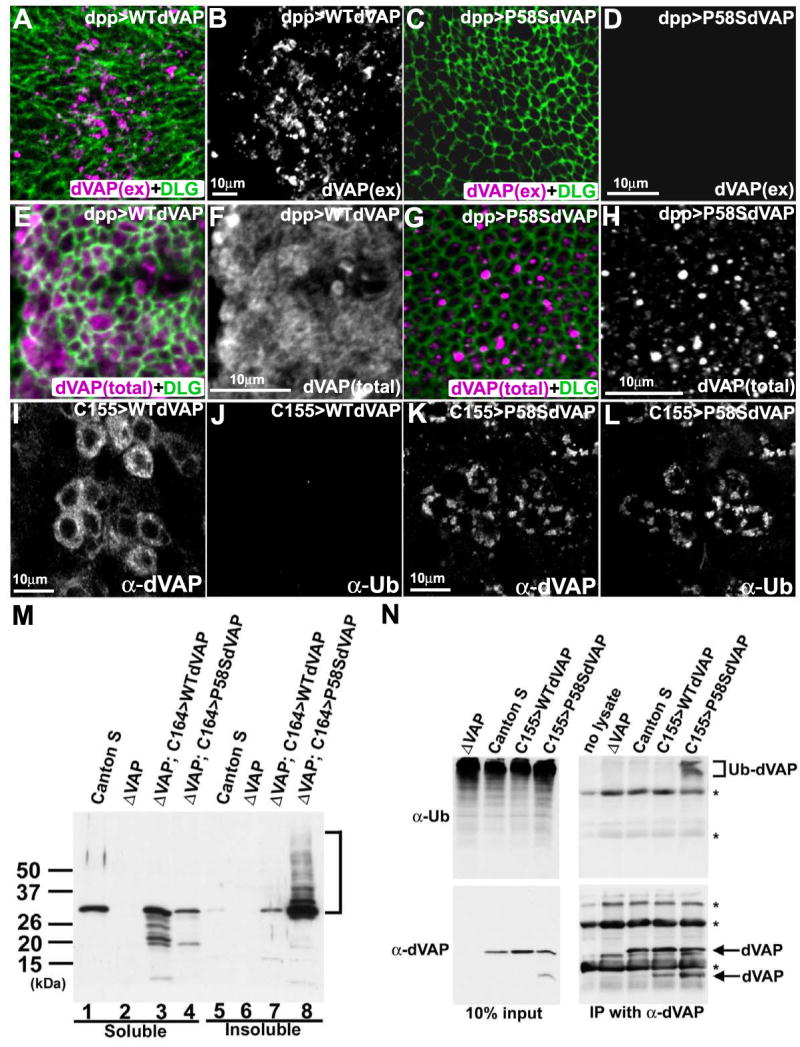
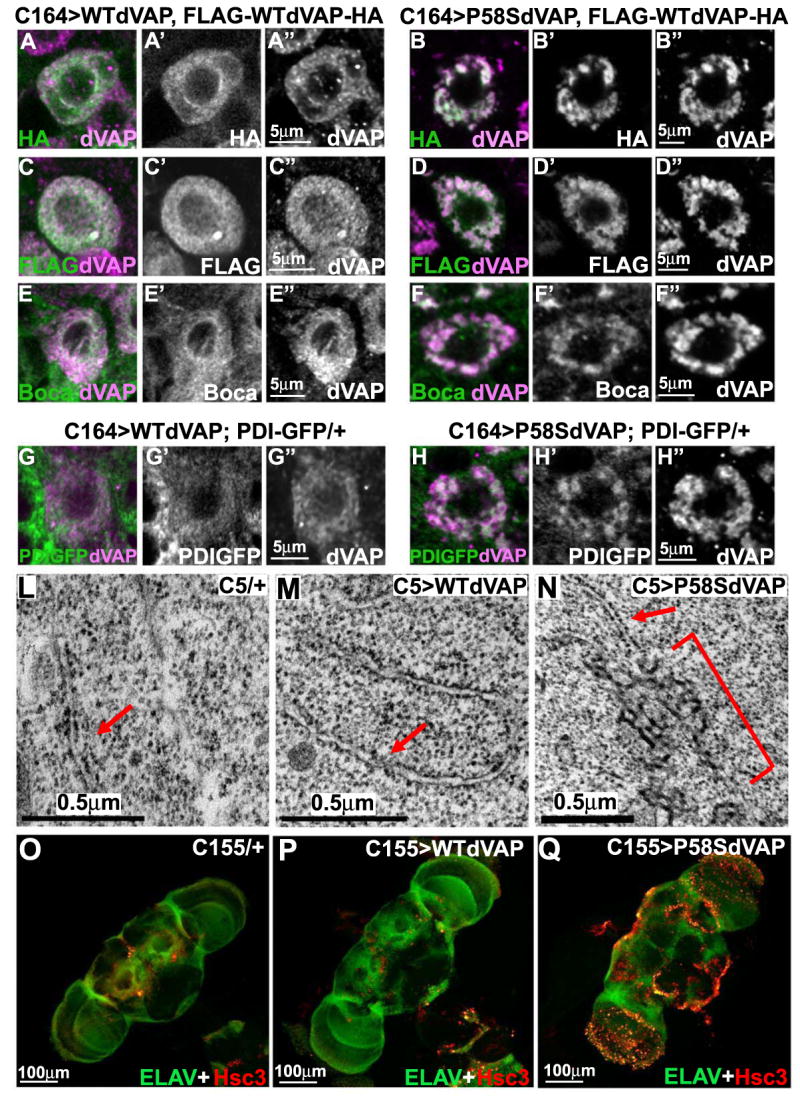
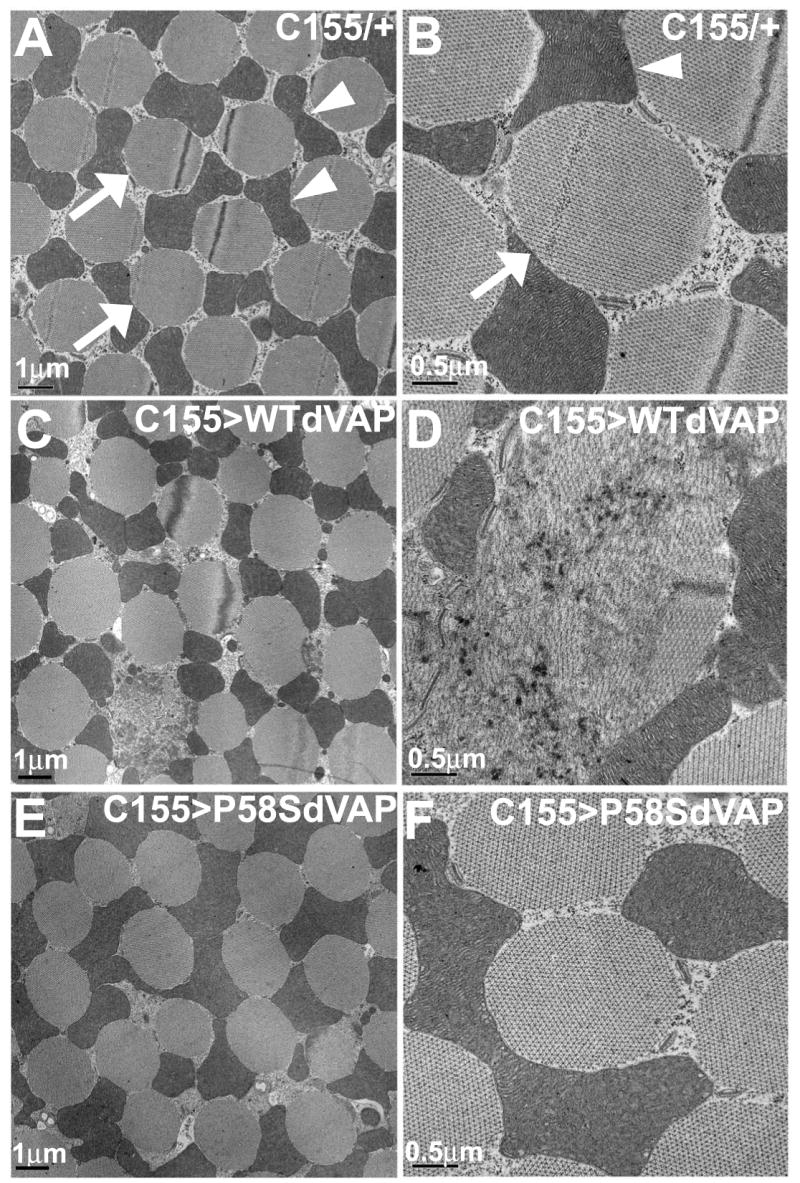
Comment in
-
From ER to Eph receptors: new roles for VAP fragments.Cell. 2008 Jun 13;133(6):949-51. doi: 10.1016/j.cell.2008.05.028. Cell. 2008. PMID: 18555770
References
-
- Amarilio R, Ramachandran S, Sabanay H, Lev S. Differential regulation of endoplasmic reticulum structure through VAP-Nir protein interaction. The Journal of biological chemistry. 2005;280:5934–5944. - PubMed
-
- Atkin JD, Farg MA, Turner BJ, Tomas D, Lysaght JA, Nunan J, Rembach A, Nagley P, Beart PM, Cheema SS, et al. Induction of the unfolded protein response in familial amyotrophic lateral sclerosis and association of protein-disulfide isomerase with superoxide dismutase 1. The Journal of biological chemistry. 2006;281:30152–30165. - PubMed
-
- Baker AM, Roberts TM, Stewart M. 2.6 A resolution crystal structure of helices of the motile major sperm protein (MSP) of Caenorhabditis elegans. J Mol Biol. 2002;319:491–499. - PubMed
-
- Basso M, Massignan T, Samengo G, Cheroni C, De Biasi S, Salmona M, Bendotti C, Bonetto V. Insoluble mutant SOD1 is partly oligoubiquitinated in amyotrophic lateral sclerosis mice. The Journal of biological chemistry. 2006;281:33325–33335. - PubMed
-
- Blelloch R, Anna-Arriola SS, Gao D, Li Y, Hodgkin J, Kimble J. The gon-1 gene is required for gonadal morphogenesis in Caenorhabditis elegans. Dev Biol. 1999;216:382–393. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
