

Rewiring the specificity of two-component signal transduction systems
- PMID: 18555780
- PMCID: PMC2453690
- DOI: 10.1016/j.cell.2008.04.040
Rewiring the specificity of two-component signal transduction systems
Abstract
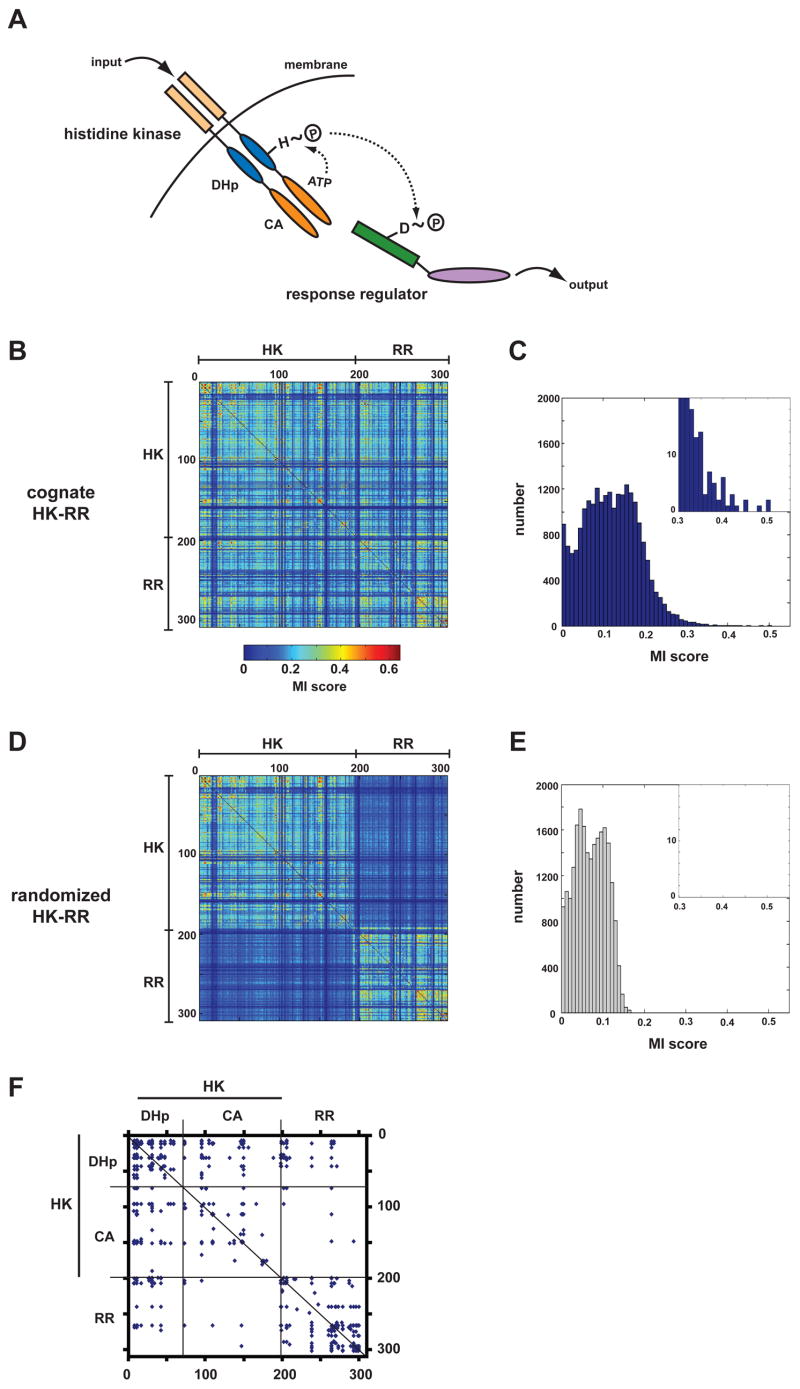
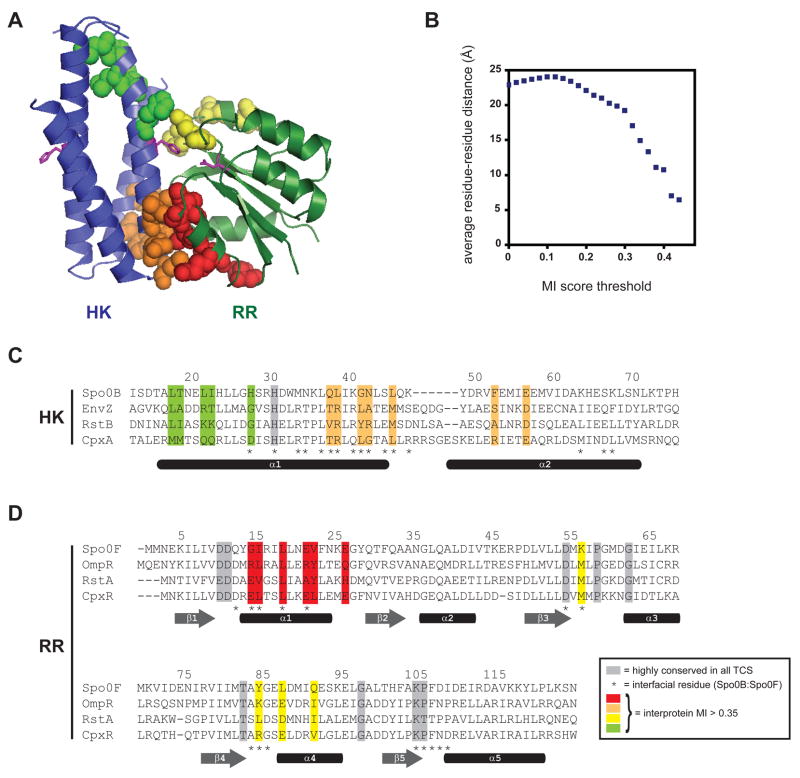
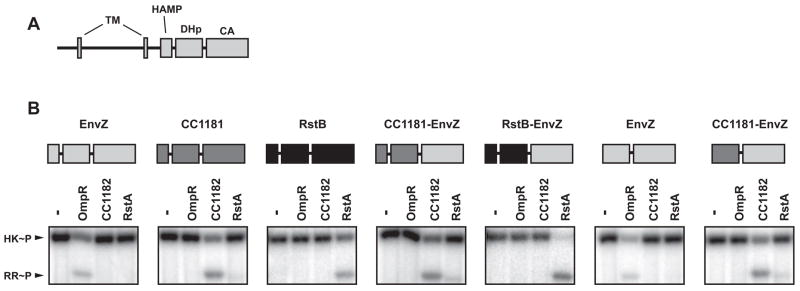
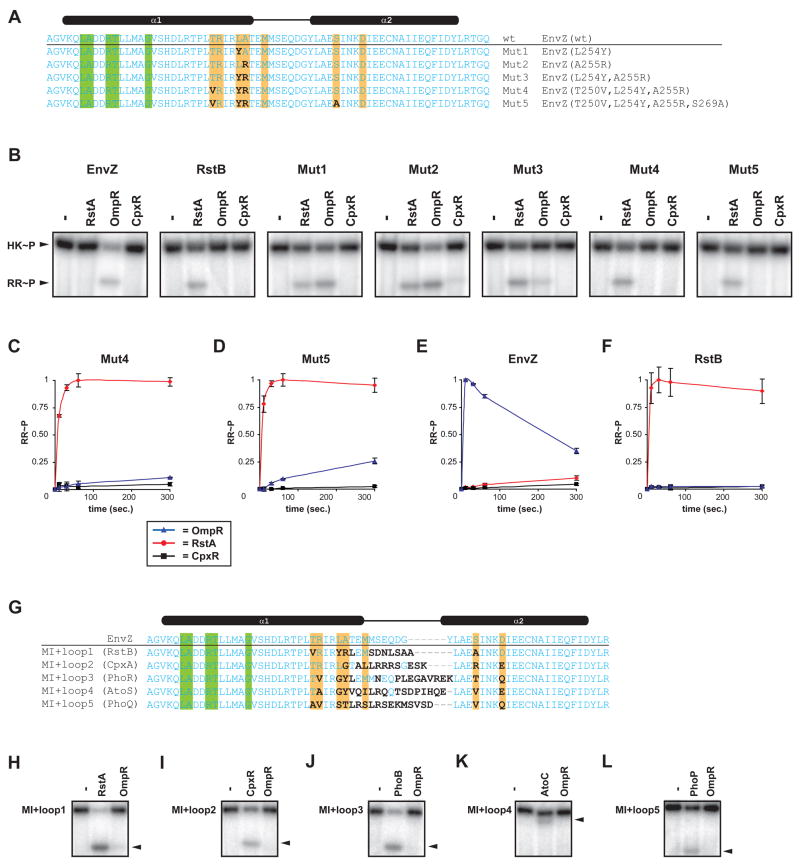
Two-component signal transduction systems are the predominant means by which bacteria sense and respond to environmental stimuli. Bacteria often employ tens or hundreds of these paralogous signaling systems, comprised of histidine kinases (HKs) and their cognate response regulators (RRs). Faithful transmission of information through these signaling pathways and avoidance of detrimental crosstalk demand exquisite specificity of HK-RR interactions. To identify the determinants of two-component signaling specificity, we examined patterns of amino acid coevolution in large, multiple sequence alignments of cognate kinase-regulator pairs. Guided by these results, we demonstrate that a subset of the coevolving residues is sufficient, when mutated, to completely switch the substrate specificity of the kinase EnvZ. Our results shed light on the basis of molecular discrimination in two-component signaling pathways, provide a general approach for the rational rewiring of these pathways, and suggest that analyses of coevolution may facilitate the reprogramming of other signaling systems and protein-protein interactions.
Figures


Comment in
-
Rewiring bacteria, two components at a time.Cell. 2008 Jun 13;133(6):947-8. doi: 10.1016/j.cell.2008.05.035. Cell. 2008. PMID: 18555769
References
-
- Atchley WR, Wollenberg KR, Fitch WM, Terhalle W, Dress AW. Correlations among amino acid sites in bHLH protein domains: an information theoretic analysis. Mol Biol Evol. 2000;17:164–178. - PubMed
-
- Bloom JD, Meyer MM, Meinhold P, Otey CR, MacMillan D, Arnold FH. Evolving strategies for enzyme engineering. Curr Opin Struct Biol. 2005;15:447–452. - PubMed
-
- Buck MJ, Atchley WR. Networks of coevolving sites in structural and functional domains of serpin proteins. Mol Biol Evol. 2005;22:1627–1634. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
