

Donor-reactive CD8 memory T cells infiltrate cardiac allografts within 24-h posttransplant in naive recipients
- PMID: 18557725
- PMCID: PMC2625311
- DOI: 10.1111/j.1600-6143.2008.02302.x
Donor-reactive CD8 memory T cells infiltrate cardiac allografts within 24-h posttransplant in naive recipients
Abstract
Normal immune responses stimulated by pathogenic and environmental antigens generate memory T cells that react with donor antigens and no currently used immunosuppressive drug completely inhibits memory T-cell function. While donor-reactive memory T cells clearly compromise graft outcomes, mechanisms utilized by memory T cells to promote rejection are largely unknown. In this study, we investigated how early endogenous memory cells infiltrate and express effector function in cardiac allografts. Endogenous CD8 memory T cells in nonsensitized recipients distinguish syngeneic versus allogeneic cardiac allografts within 24 h of reperfusion. CD8-dependent production of IFN-gamma and CXCL9/Mig was observed 24 to 72 h posttransplant in allografts but not isografts. CXCL9 was produced by donor cells in response to IFN-gamma made by recipient CD8 T cells reactive to donor class I major histocompatibility complex (MHC) molecules. Activated CD8 T cells were detected in allografts at least 3 days before donor-specific effector T cells producing IFN-gamma were detected in the recipient spleen. Early inflammation mediated by donor-reactive CD8 memory T cells greatly enhanced primed effector T-cell infiltration into allografts. These results suggest that strategies for optimal inhibition of alloimmunity should include neutralization of infiltrating CD8 memory T cells within a very narrow window after transplantation.
Figures
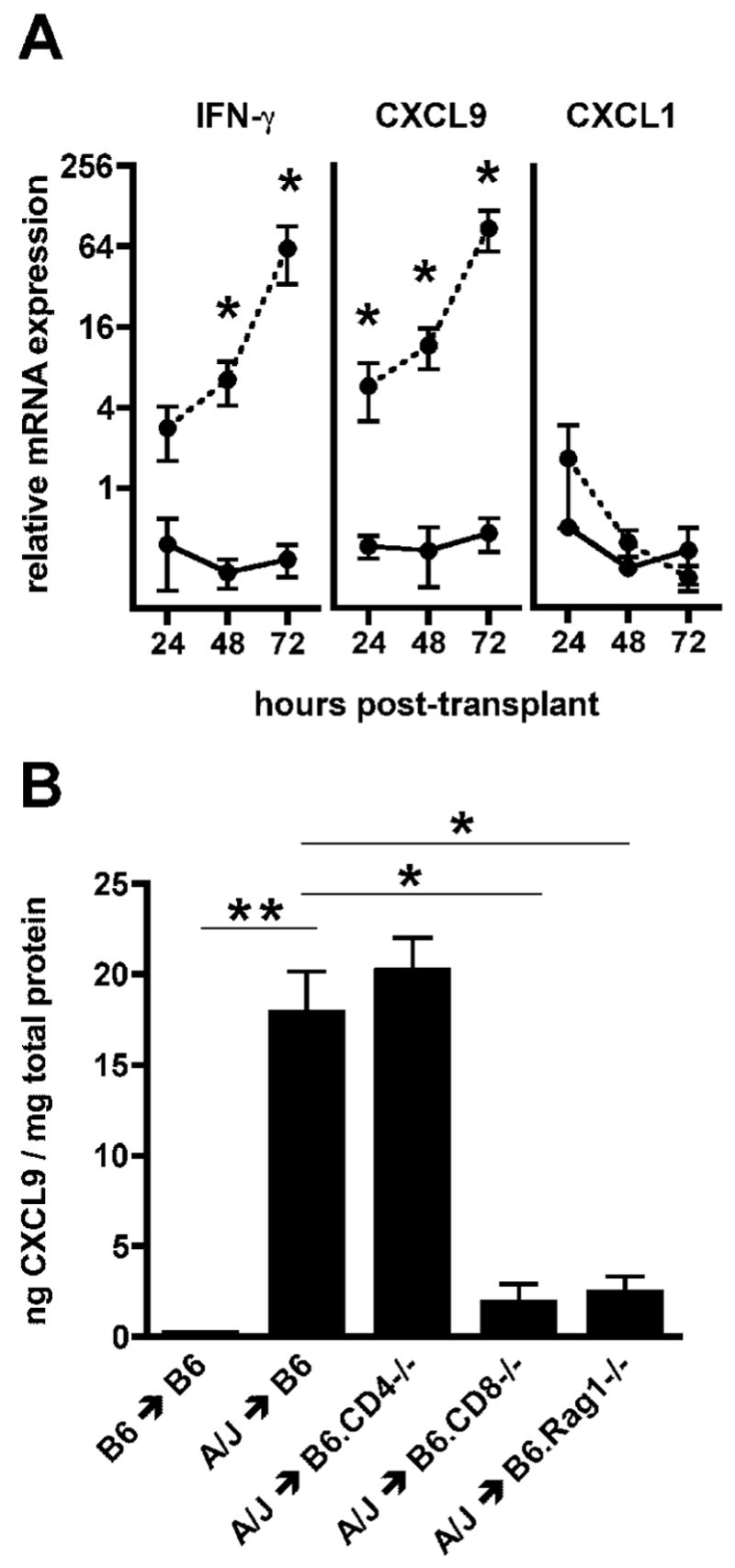
 = p ≤ 0.05) B. Wild-type C57BL/6, B6.CD4−/−, B6.CD8−/− or B6.Rag1−/− mice received syngeneic or A/J cardiac allografts. Groups of 5 grafts were harvested 72 hours post-transplant and intragraft levels of CXCL9 were measured by ELISA. (
= p ≤ 0.05) B. Wild-type C57BL/6, B6.CD4−/−, B6.CD8−/− or B6.Rag1−/− mice received syngeneic or A/J cardiac allografts. Groups of 5 grafts were harvested 72 hours post-transplant and intragraft levels of CXCL9 were measured by ELISA. (
 = p ≤ 0.0001,
= p ≤ 0.002)
= p ≤ 0.0001,
= p ≤ 0.002) = p ≤ 0.05 vs. BALB/C ≤ Rag1−/− + 2C) B. (C57BL/10xDBA/1)F1 mice received syngeneic, class II MHC-disparate bq1, or complete MHC-disparate C3H cardiac allografts. Protein levels of CXCL9 were measured in groups of 5 grafts harvested 72 hours after transplantation. (
= p ≤ 0.005 vs. complete MHC disparity)
= p ≤ 0.05 vs. BALB/C ≤ Rag1−/− + 2C) B. (C57BL/10xDBA/1)F1 mice received syngeneic, class II MHC-disparate bq1, or complete MHC-disparate C3H cardiac allografts. Protein levels of CXCL9 were measured in groups of 5 grafts harvested 72 hours after transplantation. (
= p ≤ 0.005 vs. complete MHC disparity) = p ≤ 0.01)
= p ≤ 0.01) = p ≤ 0.05,
= p ≤ 0.005)
= p ≤ 0.05,
= p ≤ 0.005) = p ≤ 0.05). C. Quantification of IFN-γ and CXCL9 mRNA within allografts 72 hours post-transplant and normalized to expression levels in the no-transfer control group (n = 4/group,
= p ≤ 0.05). D. Using the experimental design described above, 2 × 105 central memory (CD8+CD44hiCD62Lhi) or effector memory (CD8+CD44hiCD62Llo) cells were flow-sort purified and adoptively transferred into CD8−/− recipients of A/J cardiac allografts. Quantification of graft-infiltration by the adoptively transferred CD8 T cells 72 hours after transplantation is shown (n = 4/group).
= p ≤ 0.05). C. Quantification of IFN-γ and CXCL9 mRNA within allografts 72 hours post-transplant and normalized to expression levels in the no-transfer control group (n = 4/group,
= p ≤ 0.05). D. Using the experimental design described above, 2 × 105 central memory (CD8+CD44hiCD62Lhi) or effector memory (CD8+CD44hiCD62Llo) cells were flow-sort purified and adoptively transferred into CD8−/− recipients of A/J cardiac allografts. Quantification of graft-infiltration by the adoptively transferred CD8 T cells 72 hours after transplantation is shown (n = 4/group). = p ≤ 0.05 vs. IgG).
= p ≤ 0.05 vs. IgG).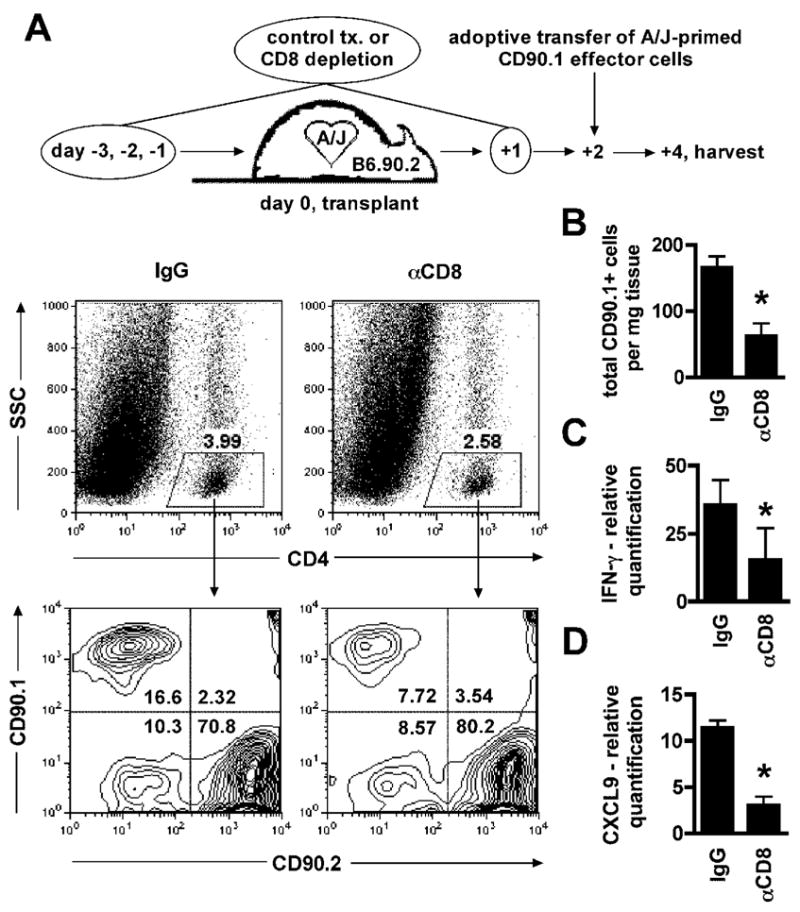 = p ≤ 0.05). C–D. Relative mRNA quantification of IFN-γ and CXCL9 in grafts harvested from IgG and γ-CD8 treated recipients 4 days after transplantation and 48 hours after adoptive transfer. Data are normalized to a sample randomly chosen from the γ-CD8 treated group (n = 6/group,
= p ≤ 0.05).
= p ≤ 0.05). C–D. Relative mRNA quantification of IFN-γ and CXCL9 in grafts harvested from IgG and γ-CD8 treated recipients 4 days after transplantation and 48 hours after adoptive transfer. Data are normalized to a sample randomly chosen from the γ-CD8 treated group (n = 6/group,
= p ≤ 0.05).References
-
- Lechler RI, Sykes M, Thomson AW, Turka LA. Organ transplantation--how much of the promise has been realized? Nat Med. 2005;11(6):605–613. - PubMed
-
- Fishman JA. Infection in solid-organ transplant recipients. N Engl J Med. 2007;357(25):2601–2614. - PubMed
-
- Ajithkumar TV, Parkinson CA, Butler A, Hatcher HM. Management of solid tumours in organ-transplant recipients. Lancet Oncol. 2007;8(10):921–932. - PubMed
-
- Sayegh MH, Remuzzi G. Clinical update: immunosuppression minimisation. Lancet. 2007;369(9574):1676–1678. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
