

Redundant synthesis of cysteinyl-tRNACys in Methanosarcina mazei
- PMID: 18559341
- PMCID: PMC2494925
- DOI: 10.1074/jbc.M801839200
Redundant synthesis of cysteinyl-tRNACys in Methanosarcina mazei
Abstract
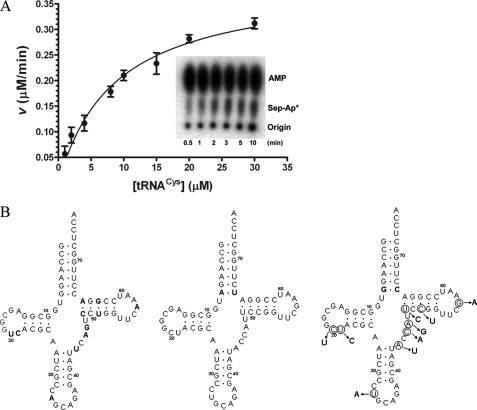
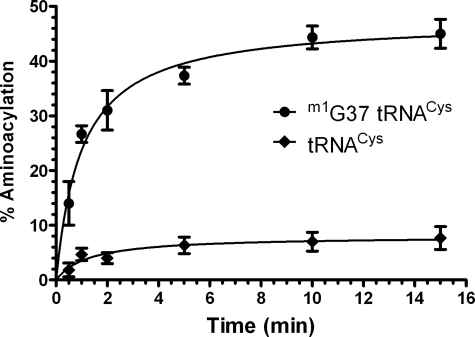
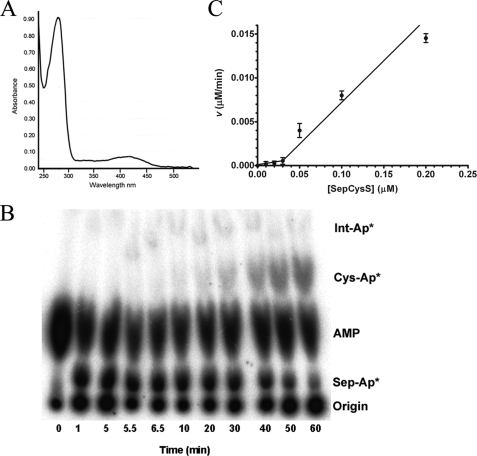
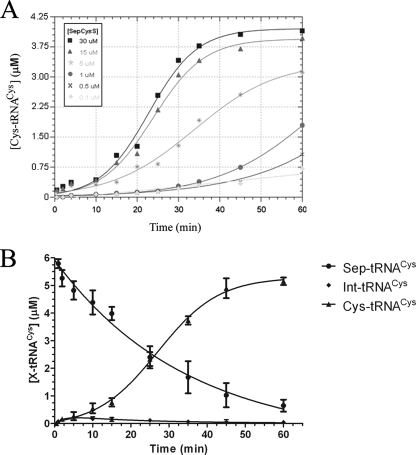
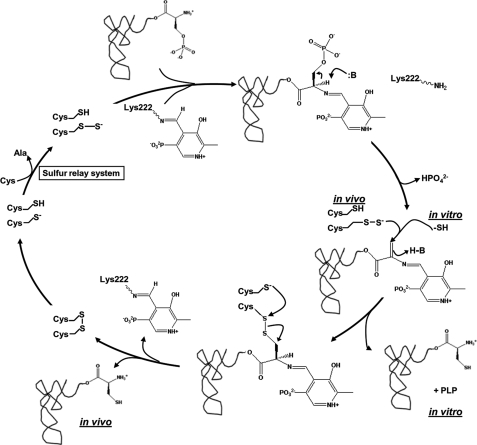
A subset of methanogenic archaea synthesize the cysteinyl-tRNA(Cys) (Cys-tRNA(Cys)) needed for protein synthesis using both a canonical cysteinyl-tRNA synthetase (CysRS) as well as a set of two enzymes that operate via a separate indirect pathway. In the indirect route, phosphoseryl-tRNA(Cys) (Sep-tRNA(Cys)) is first synthesized by phosphoseryl-tRNA synthetase (SepRS), and this misacylated intermediate is then converted to Cys-tRNA(Cys) by Sep-tRNA:Cys-tRNA synthase (SepCysS) via a pyridoxal phosphate-dependent mechanism. Here, we explore the function of all three enzymes in the mesophilic methanogen Methanosarcina mazei. The genome of M. mazei also features three distinct tRNA(Cys) isoacceptors, further indicating the unusual and complex nature of Cys-tRNA(Cys) synthesis in this organism. Comparative aminoacylation kinetics by M. mazei CysRS and SepRS reveals that each enzyme prefers a distinct tRNA(Cys) isoacceptor or pair of isoacceptors. Recognition determinants distinguishing the tRNAs are shown to reside in the globular core of the molecule. Both enzymes also require the S-adenosylmethione-dependent formation of (m1)G37 in the anticodon loop for efficient aminoacylation. We further report a new, highly sensitive assay to measure the activity of SepCysS under anaerobic conditions. With this approach, we demonstrate that SepCysS functions as a multiple-turnover catalyst with kinetic behavior similar to bacterial selenocysteine synthase and the archaeal/eukaryotic SepSecS enzyme. Together, these data suggest that both metabolic routes and all three tRNA(Cys) species in M. mazei play important roles in the cellular physiology of the organism.
Figures
Similar articles
-
Aminoacylation of tRNA with phosphoserine for synthesis of cysteinyl-tRNA(Cys).Nat Struct Mol Biol. 2008 May;15(5):507-14. doi: 10.1038/nsmb.1423. Epub 2008 Apr 20. Nat Struct Mol Biol. 2008. PMID: 18425141 Free PMC article.
-
RNA-Dependent Cysteine Biosynthesis in Bacteria and Archaea.mBio. 2017 May 9;8(3):e00561-17. doi: 10.1128/mBio.00561-17. mBio. 2017. PMID: 28487430 Free PMC article.
-
The homotetrameric phosphoseryl-tRNA synthetase from Methanosarcina mazei exhibits half-of-the-sites activity.J Biol Chem. 2008 Aug 8;283(32):21997-2006. doi: 10.1074/jbc.M801838200. Epub 2008 Jun 17. J Biol Chem. 2008. PMID: 18559342 Free PMC article.
-
From one amino acid to another: tRNA-dependent amino acid biosynthesis.Nucleic Acids Res. 2008 Apr;36(6):1813-25. doi: 10.1093/nar/gkn015. Epub 2008 Feb 5. Nucleic Acids Res. 2008. PMID: 18252769 Free PMC article. Review.
-
Cys-tRNACys formation and cysteine biosynthesis in methanogenic archaea: two faces of the same problem?Cell Mol Life Sci. 2004 Oct;61(19-20):2437-45. doi: 10.1007/s00018-004-4194-9. Cell Mol Life Sci. 2004. PMID: 15526152 Free PMC article. Review.
Cited by
-
Mechanism of N-methylation by the tRNA m1G37 methyltransferase Trm5.RNA. 2010 Dec;16(12):2484-92. doi: 10.1261/rna.2376210. Epub 2010 Oct 27. RNA. 2010. PMID: 20980671 Free PMC article.
-
Machine Learning Algorithms Highlight tRNA Information Content and Chargaff's Second Parity Rule Score as Important Features in Discriminating Probiotics from Non-Probiotics.Biology (Basel). 2022 Jul 7;11(7):1024. doi: 10.3390/biology11071024. Biology (Basel). 2022. PMID: 36101405 Free PMC article.
-
Specific and nonhomologous isofunctional enzymes of the genetic information processing pathways as potential therapeutical targets for tritryps.Enzyme Res. 2011;2011:543912. doi: 10.4061/2011/543912. Epub 2011 Jul 26. Enzyme Res. 2011. PMID: 21808726 Free PMC article.
-
tRNAs: cellular barcodes for amino acids.FEBS Lett. 2010 Jan 21;584(2):387-95. doi: 10.1016/j.febslet.2009.11.013. FEBS Lett. 2010. PMID: 19903480 Free PMC article. Review.
-
Efficient Sulfide Assimilation in Methanosarcina acetivorans Is Mediated by the MA1715 Protein.J Bacteriol. 2016 Jun 27;198(14):1974-83. doi: 10.1128/JB.00141-16. Print 2016 Jul 15. J Bacteriol. 2016. PMID: 27137504 Free PMC article.
References
-
- Deppenmeier, U., Johann, A., Hartsch, T., Merkl, R., Schmitz, R. A., Martinez-Arias, R., Henne, A., Wiezer, A., Bäumer, S., Jacobi, C., Brüggemann, H., Lienard, T., Christmann, A., Bömeke, M., Steckel, S., Bhattacharyya, A., Lykidis, A., Overbeek, R., Klenk, H.-P., Gunsalus, R. P., Fritz, H.-J., and Gottschalk, G. (2002) J. Mol. Microbiol. Biotechnol. 4 453-461 - PubMed
-
- Boetius, A., Ravenschlag, K., Schubert, C. J., Rickert, D., Widdel, F., Gleseke, A., Amann, R., Jergensen, B. B., Witte, U., and Pfannkuche, O. (2000) Nature 407 623-626 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
