

Curvature and torsion in growing actin networks
- PMID: 18560043
- PMCID: PMC3236663
- DOI: 10.1088/1478-3975/5/2/026006
Curvature and torsion in growing actin networks
Abstract
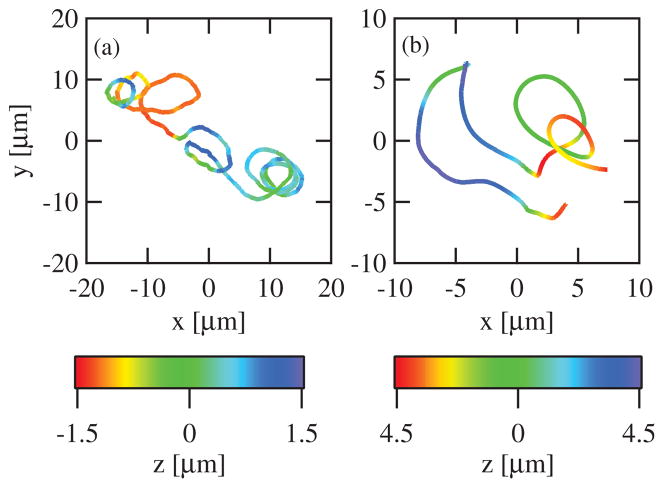
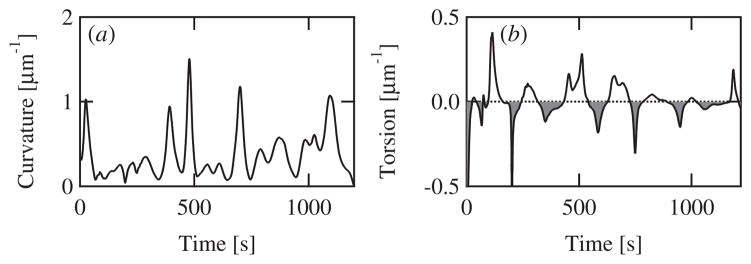
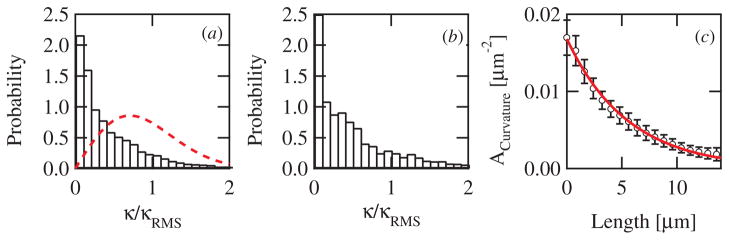
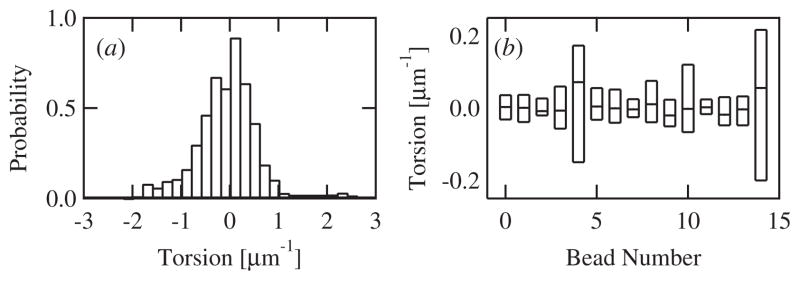
Intracellular pathogens such as Listeria monocytogenes and Rickettsia rickettsii move within a host cell by polymerizing a comet-tail of actin fibers that ultimately pushes the cell forward. This dense network of cross-linked actin polymers typically exhibits a striking curvature that causes bacteria to move in gently looping paths. Theoretically, tail curvature has been linked to details of motility by considering force and torque balances from a finite number of polymerizing filaments. Here we track beads coated with a prokaryotic activator of actin polymerization in three dimensions to directly quantify the curvature and torsion of bead motility paths. We find that bead paths are more likely to have low rather than high curvature at any given time. Furthermore, path curvature changes very slowly in time, with an autocorrelation decay time of 200 s. Paths with a small radius of curvature, therefore, remain so for an extended period resulting in loops when confined to two dimensions. When allowed to explore a three-dimensional (3D) space, path loops are less evident. Finally, we quantify the torsion in the bead paths and show that beads do not exhibit a significant left- or right-handed bias to their motion in 3D. These results suggest that paths of actin-propelled objects may be attributed to slow changes in curvature, possibly associated with filament debranching, rather than a fixed torque.
Figures
Similar articles
-
Dynamics of actin-based movement by Rickettsia rickettsii in vero cells.Infect Immun. 1999 Aug;67(8):4201-7. doi: 10.1128/IAI.67.8.4201-4207.1999. Infect Immun. 1999. PMID: 10417192 Free PMC article.
-
Curved tails in polymerization-based bacterial motility.Phys Rev E Stat Nonlin Soft Matter Phys. 2001 Aug;64(2 Pt 1):021904. doi: 10.1103/PhysRevE.64.021904. Epub 2001 Jul 19. Phys Rev E Stat Nonlin Soft Matter Phys. 2001. PMID: 11497617
-
A comparative study of the actin-based motilities of the pathogenic bacteria Listeria monocytogenes, Shigella flexneri and Rickettsia conorii.J Cell Sci. 1999 Jun;112 ( Pt 11):1697-708. doi: 10.1242/jcs.112.11.1697. J Cell Sci. 1999. PMID: 10318762
-
The actin-based motility of the facultative intracellular pathogen Listeria monocytogenes.Mol Microbiol. 1994 Aug;13(3):395-402. doi: 10.1111/j.1365-2958.1994.tb00434.x. Mol Microbiol. 1994. PMID: 7997157 Review.
-
Biomimetic systems for studying actin-based motility.Curr Biol. 2003 Sep 16;13(18):R734-44. doi: 10.1016/j.cub.2003.08.051. Curr Biol. 2003. PMID: 13678615 Review.
Cited by
-
Mesoscopic model of actin-based propulsion.PLoS Comput Biol. 2012;8(11):e1002764. doi: 10.1371/journal.pcbi.1002764. Epub 2012 Nov 1. PLoS Comput Biol. 2012. PMID: 23133366 Free PMC article.
-
Observation and kinematic description of long actin tracks induced by spherical beads.Biophys J. 2010 Nov 3;99(9):2793-802. doi: 10.1016/j.bpj.2010.08.058. Biophys J. 2010. PMID: 21044576 Free PMC article.
References
-
- Welch MD, Mullins RD. Cellular control of actin nucleation. Annu Rev Cell Dev Biol. 2002;18:247–88. - PubMed
-
- Pollard TD, Borisy G. Cellular motility driven by assembly and disassembly of actin filaments. Cell. 2003;112:453–65. - PubMed
-
- Fletcher DA, Theriot JA. An introduction to cell motility for the physical scientist. Phys Biol. 2004;1:T1–10. - PubMed
-
- Gerbal F, Laurent V, Ott A, Carlier MF, Chaikin P, Prost J. Measurement of the elasticity of the actin tail of listeria monocytogenes. Eur Biophys J Biophys Lett. 2000;29:134–40. - PubMed
-
- Jeng RL, Goley ED, D’Alessio JA, Chaga OY, Svitkina TM, Borisy GG, Heinzen RA, Welch MD. A rickettsia wasp-like protein activates the arp2/3 complex and mediates actin-based motility. Cell Microbiol. 2004;6:761–9. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources