

Effects of nerve injury and segmental regeneration on the cellular correlates of neural morphallaxis
- PMID: 18561185
- PMCID: PMC2754161
- DOI: 10.1002/jez.b.21224
Effects of nerve injury and segmental regeneration on the cellular correlates of neural morphallaxis
Abstract
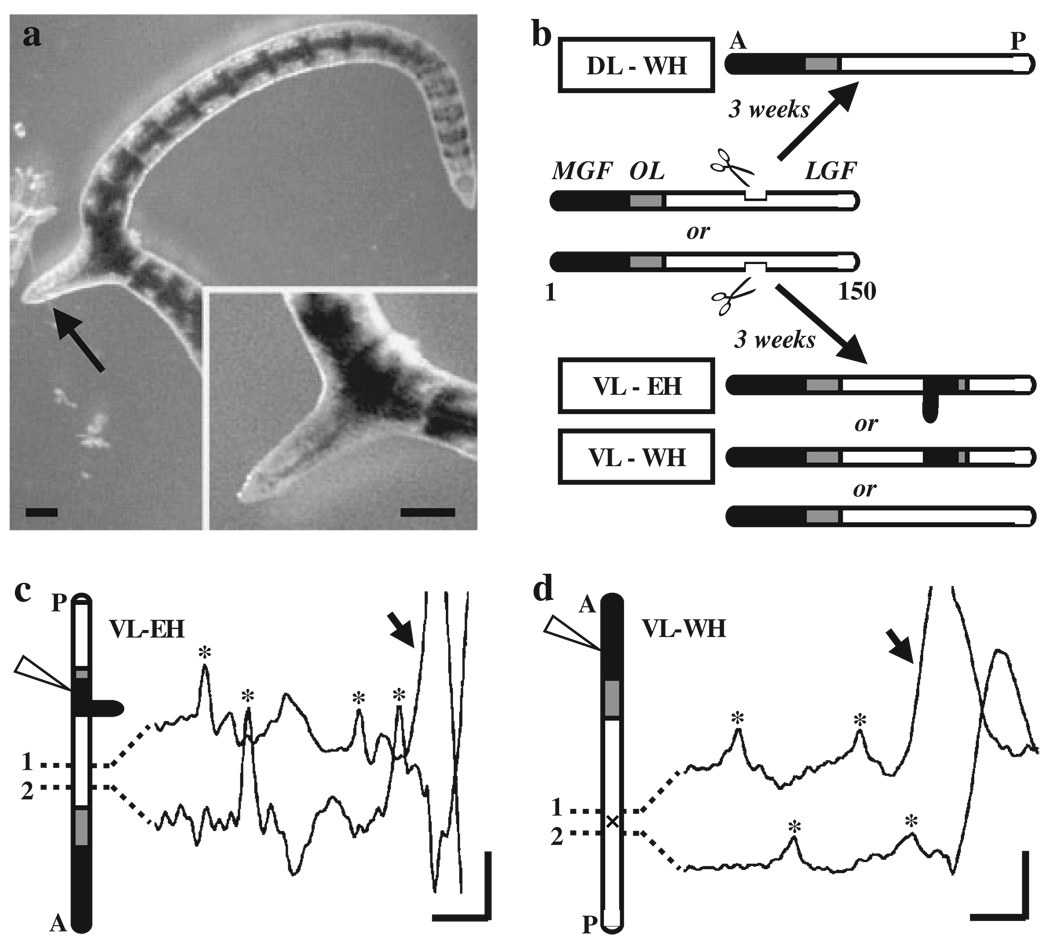
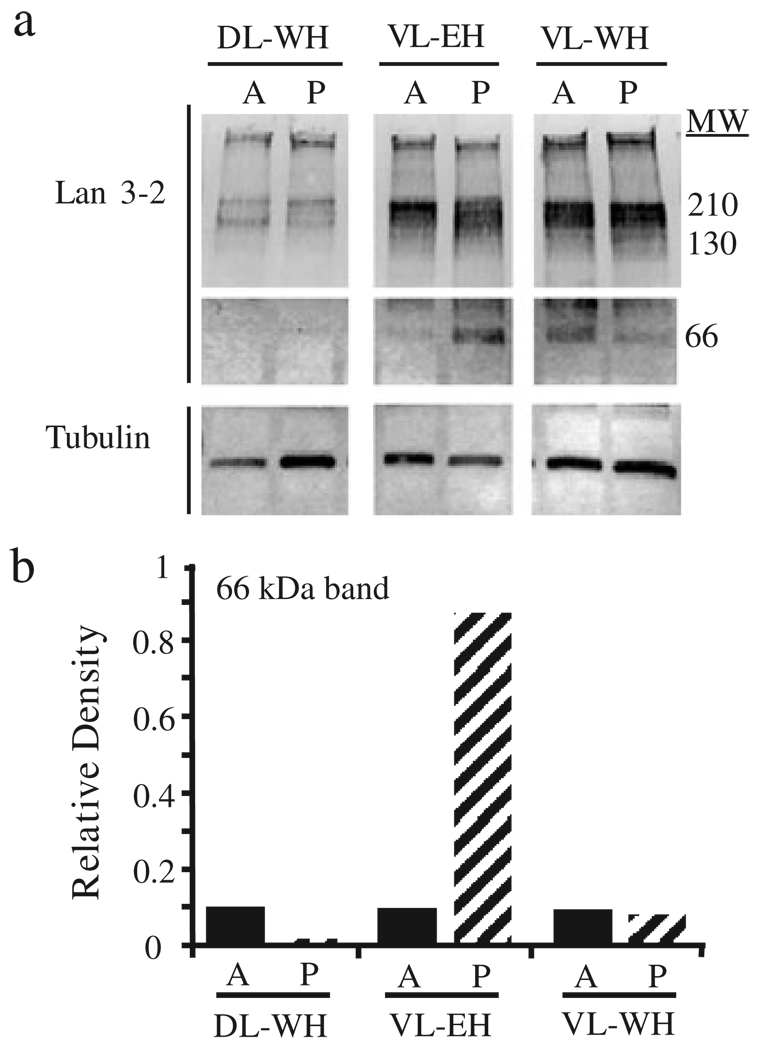
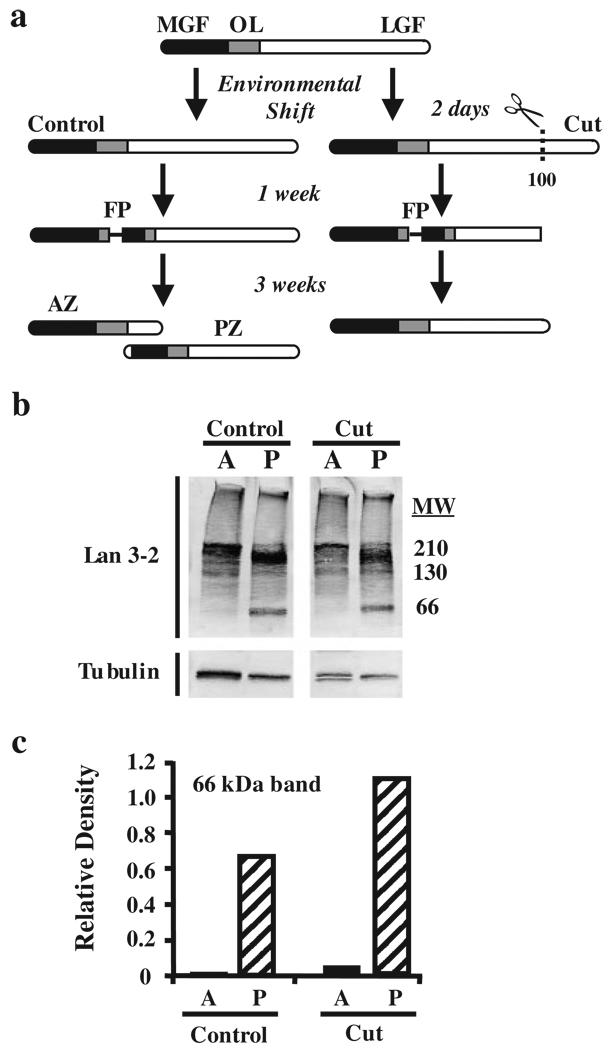
Functional recovery of neural networks after injury requires a series of signaling events similar to the embryonic processes that governed initial network construction. Neural morphallaxis, a form of nervous system regeneration, involves reorganization of adult neural connectivity patterns. Neural morphallaxis in the worm, Lumbriculus variegatus, occurs during asexual reproduction and segmental regeneration, as body fragments acquire new positional identities along the anterior-posterior axis. Ectopic head (EH) formation, induced by ventral nerve cord lesion, generated morphallactic plasticity including the reorganization of interneuronal sensory fields and the induction of a molecular marker of neural morphallaxis. Morphallactic changes occurred only in segments posterior to an EH. Neither EH formation, nor neural morphallaxis was observed after dorsal body lesions, indicating a role for nerve cord injury in morphallaxis induction. Furthermore, a hierarchical system of neurobehavioral control was observed, where anterior heads were dominant and an EH controlled body movements only in the absence of the anterior head. Both suppression of segmental regeneration and blockade of asexual fission, after treatment with boric acid, disrupted the maintenance of neural morphallaxis, but did not block its induction. Therefore, segmental regeneration (i.e., epimorphosis) may not be required for the induction of morphallactic remodeling of neural networks. However, on-going epimorphosis appears necessary for the long-term consolidation of cellular and molecular mechanisms underlying the morphallaxis of neural circuitry.
Figures



Similar articles
-
Regeneration and asexual reproduction share common molecular changes: upregulation of a neural glycoepitope during morphallaxis in Lumbriculus.Mech Dev. 2005 May;122(5):721-32. doi: 10.1016/j.mod.2004.12.003. Epub 2004 Dec 27. Mech Dev. 2005. PMID: 15817228
-
Rapid neural circuit switching mediated by synaptic plasticity during neural morphallactic regeneration.Dev Neurobiol. 2012 Sep;72(9):1256-66. doi: 10.1002/dneu.20993. Epub 2012 Jun 21. Dev Neurobiol. 2012. PMID: 22021133
-
Morphallaxis in an aquatic oligochaete, Lumbriculus variegatus: reorganization of escape reflexes in regenerating body fragments.Dev Biol. 1990 Mar;138(1):94-103. doi: 10.1016/0012-1606(90)90179-m. Dev Biol. 1990. PMID: 2307291
-
Transplants and neurotrophic factors increase regeneration and recovery of function after spinal cord injury.Prog Brain Res. 2002;137:257-73. doi: 10.1016/s0079-6123(02)37020-1. Prog Brain Res. 2002. PMID: 12440372 Review.
-
Regeneration mechanisms in Syllidae (Annelida).Regeneration (Oxf). 2018 Apr 14;5(1):26-42. doi: 10.1002/reg2.98. eCollection 2018 Mar. Regeneration (Oxf). 2018. PMID: 29721325 Free PMC article. Review.
Cited by
-
Annotation of nerve cord transcriptome in earthworm Eisenia fetida.Genom Data. 2017 Oct 12;14:91-105. doi: 10.1016/j.gdata.2017.10.002. eCollection 2017 Dec. Genom Data. 2017. PMID: 29204349 Free PMC article.
-
It Cuts Both Ways: An Annelid Model System for the Study of Regeneration in the Laboratory and in the Classroom.Front Cell Dev Biol. 2021 Nov 29;9:780422. doi: 10.3389/fcell.2021.780422. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 34912808 Free PMC article. Review.
-
A Stable Thoracic Hox Code and Epimorphosis Characterize Posterior Regeneration in Capitella teleta.PLoS One. 2016 Feb 19;11(2):e0149724. doi: 10.1371/journal.pone.0149724. eCollection 2016. PLoS One. 2016. PMID: 26894631 Free PMC article.
-
Comparative Aspects of Annelid Regeneration: Towards Understanding the Mechanisms of Regeneration.Genes (Basel). 2021 Jul 28;12(8):1148. doi: 10.3390/genes12081148. Genes (Basel). 2021. PMID: 34440322 Free PMC article. Review.
References
-
- Achee NL, Zoran MJ. Short-term and long-term alterations in neuronal excitability during injury-induced axonal regeneration in ganglia and cell culture. Invert Neurosci. 1996;2:189–198.
-
- Agata K, Saito Y, Nakajima E. Unifying principles of regeneration I: epimorphosis versus morphallaxis. Dev Growth Differ. 2007;49:73–78. - PubMed
-
- Ambron RT, Walters ET. Priming events and retrograde injury signals. Mol Neurobiol. 1996;13:61–79. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
