

Endocannabinoid signaling controls pyramidal cell specification and long-range axon patterning
- PMID: 18562289
- PMCID: PMC2438381
- DOI: 10.1073/pnas.0803545105
Endocannabinoid signaling controls pyramidal cell specification and long-range axon patterning
Abstract
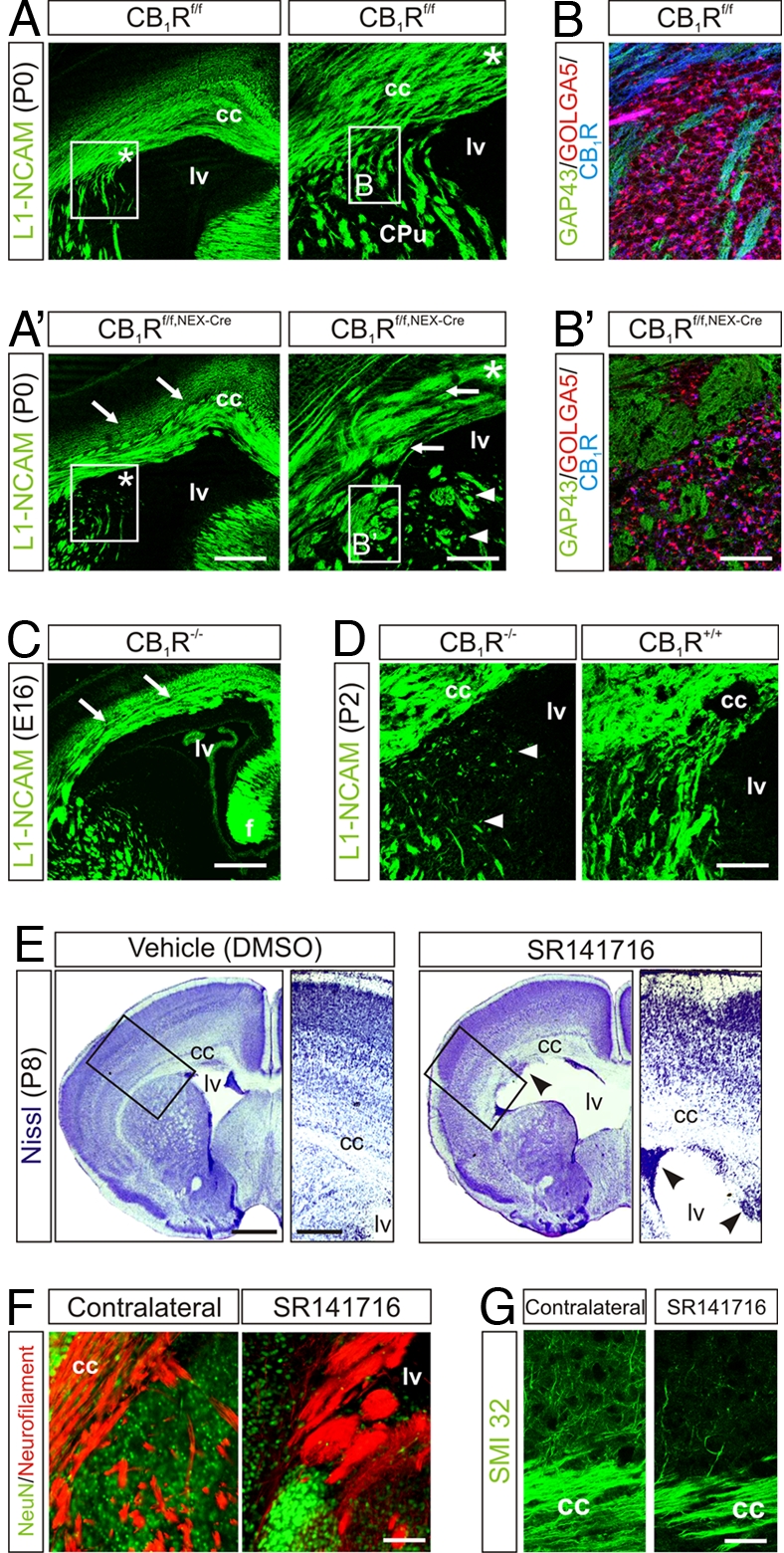
Endocannabinoids (eCBs) have recently been identified as axon guidance cues shaping the connectivity of local GABAergic interneurons in the developing cerebrum. However, eCB functions during pyramidal cell specification and establishment of long-range axonal connections are unknown. Here, we show that eCB signaling is operational in subcortical proliferative zones from embryonic day 12 in the mouse telencephalon and controls the proliferation of pyramidal cell progenitors and radial migration of immature pyramidal cells. When layer patterning is accomplished, developing pyramidal cells rely on eCB signaling to initiate the elongation and fasciculation of their long-range axons. Accordingly, CB(1) cannabinoid receptor (CB(1)R) null and pyramidal cell-specific conditional mutant (CB(1)R(f/f,NEX-Cre)) mice develop deficits in neuronal progenitor proliferation and axon fasciculation. Likewise, axonal pathfinding becomes impaired after in utero pharmacological blockade of CB(1)Rs. Overall, eCBs are fundamental developmental cues controlling pyramidal cell development during corticogenesis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
- R01DA023214/DA/NIDA NIH HHS/United States
- MC_U117570528/MRC_/Medical Research Council/United Kingdom
- R01 NS048884/NS/NINDS NIH HHS/United States
- T32 ES007332/ES/NIEHS NIH HHS/United States
- R01 DA011322/DA/NIDA NIH HHS/United States
- NS048884/NS/NINDS NIH HHS/United States
- DA21696/DA/NIDA NIH HHS/United States
- DA15916/DA/NIDA NIH HHS/United States
- K05 DA021696/DA/NIDA NIH HHS/United States
- DA11322/DA/NIDA NIH HHS/United States
- ES07332/ES/NIEHS NIH HHS/United States
- R01 DA023214/DA/NIDA NIH HHS/United States
- P01 DA015916/DA/NIDA NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
