

Serotonin evokes endocannabinoid release and retrogradely suppresses excitatory synapses
- PMID: 18562622
- PMCID: PMC2720684
- DOI: 10.1523/JNEUROSCI.0678-08.2008
Serotonin evokes endocannabinoid release and retrogradely suppresses excitatory synapses
Abstract
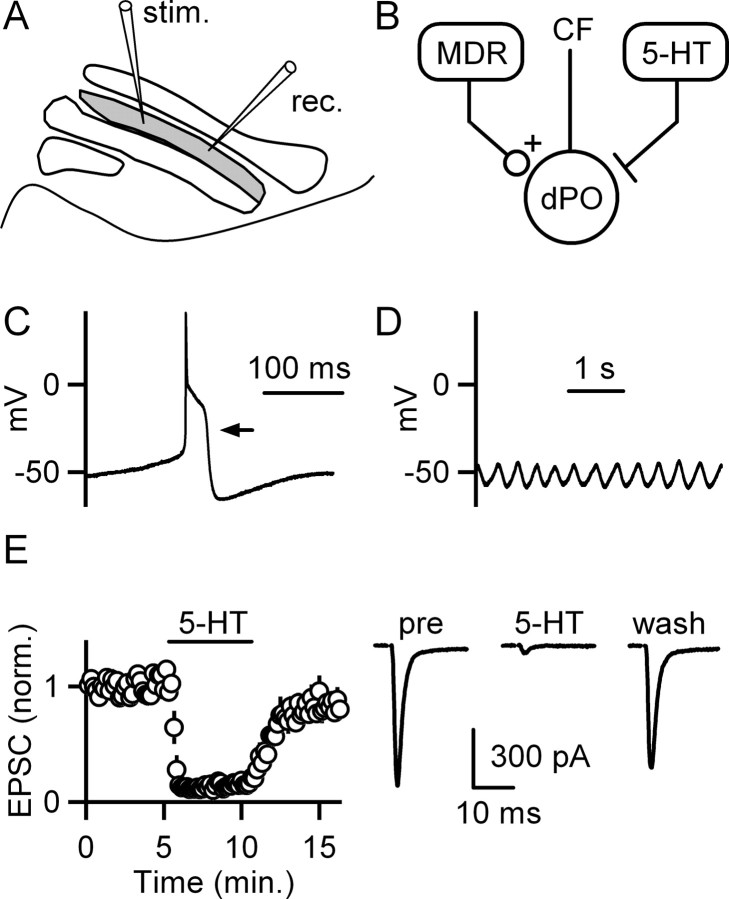
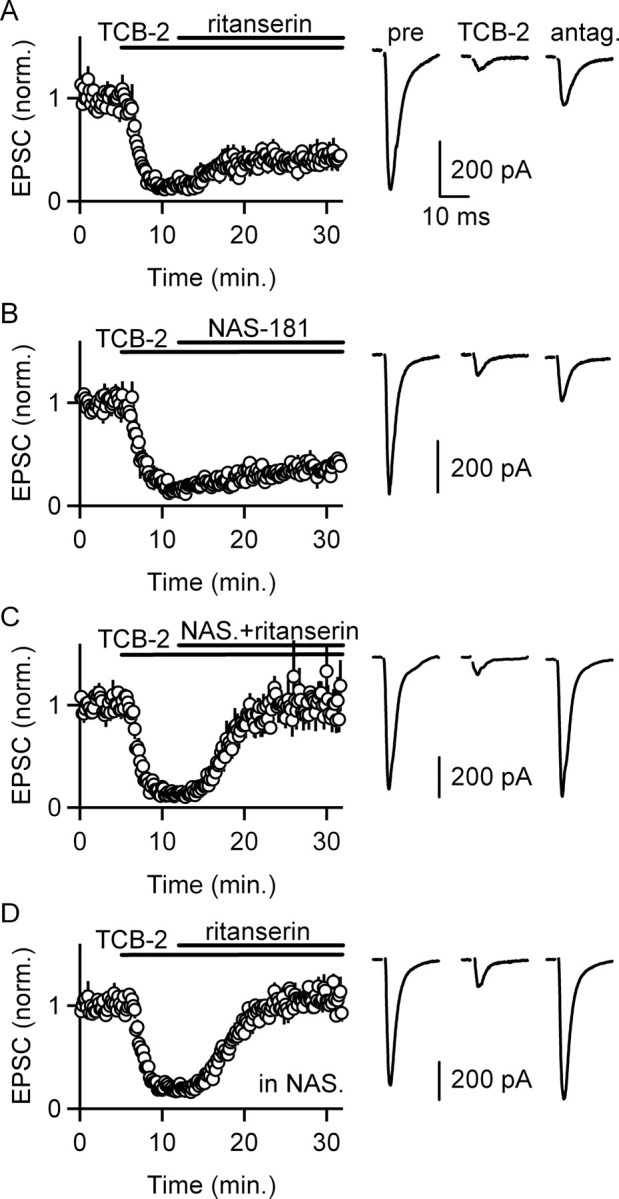
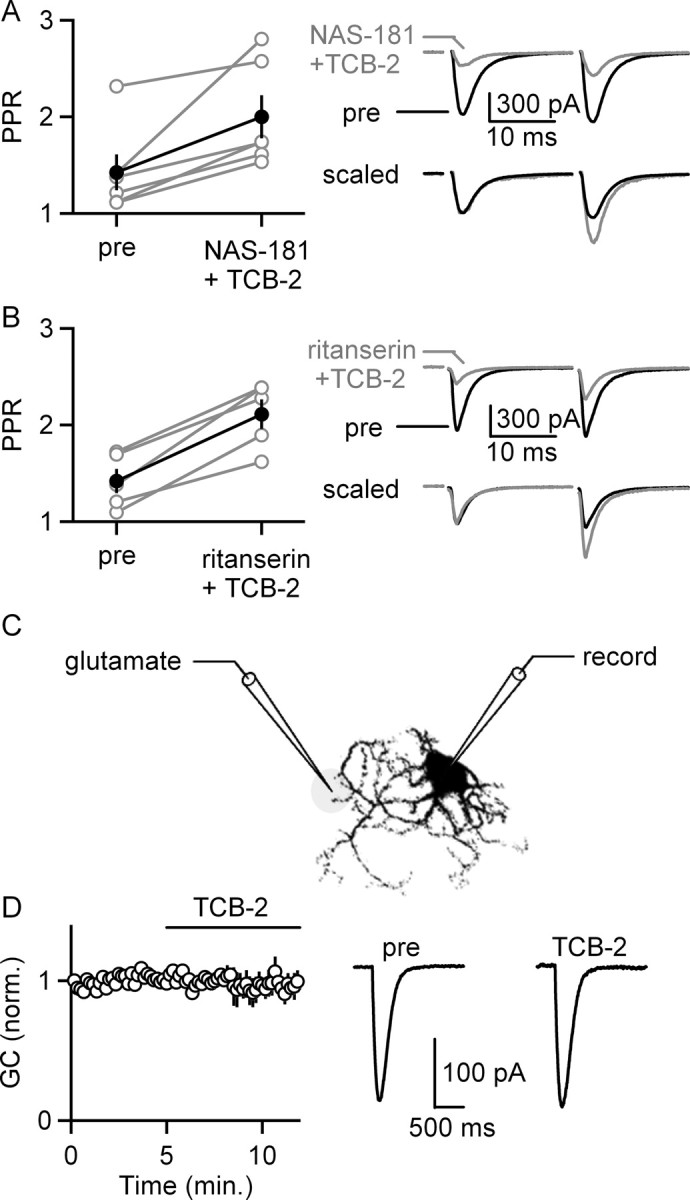
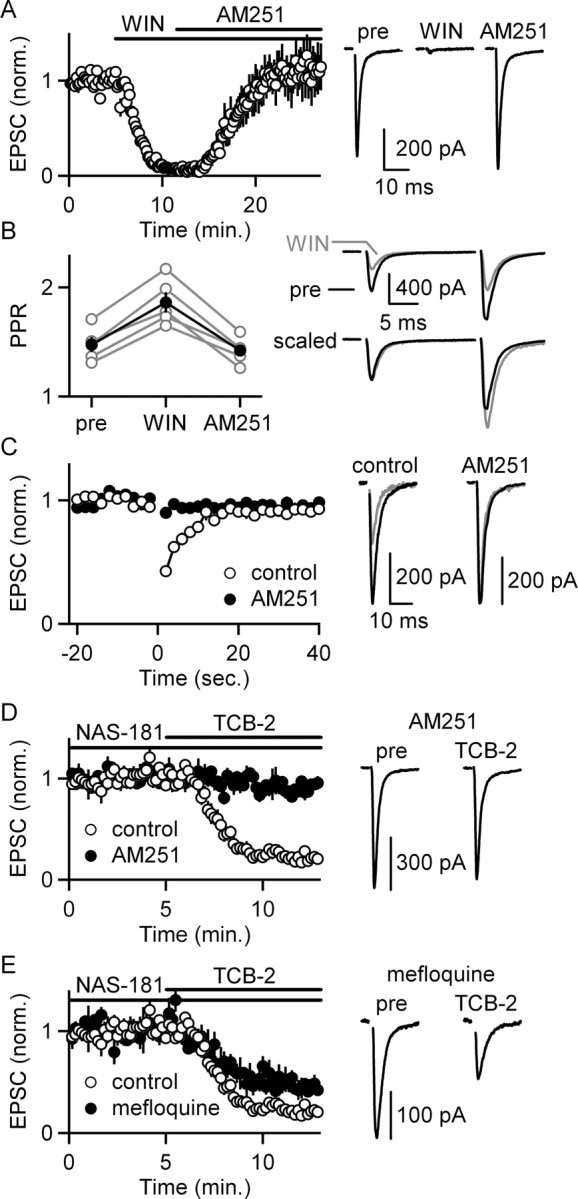
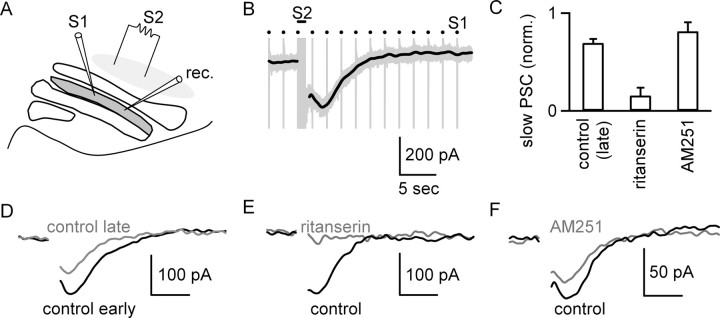
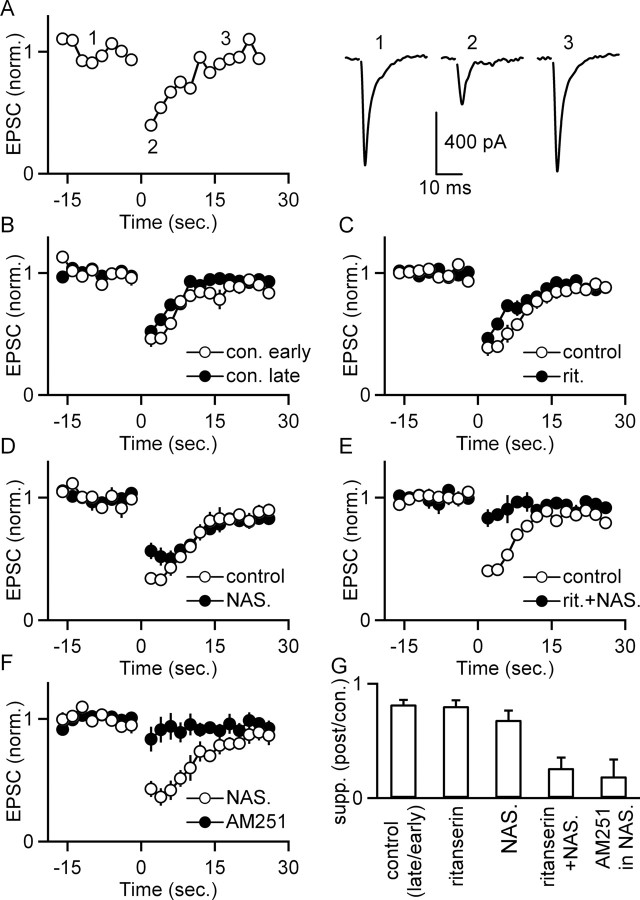
5-HT(2)-type serotonin receptors (5-HT(2)Rs) are widely expressed throughout the brain and mediate many of the modulatory effects of serotonin. It has been thought that postsynaptic 5-HT(2)Rs act primarily by depolarizing neurons and thereby increasing their excitability. However, it is also known that 5-HT(2)Rs are coupled to G(q/11)-type G-proteins and that some other types of G(q/11)-coupled receptors can regulate synapses by evoking endocannabinoid release and activating presynaptic cannabinoid-type 1 receptors (CB(1)Rs). Here, we examine whether activation of 5-HT(2)Rs can regulate synapses through such a mechanism by studying excitatory synapses onto cells in the inferior olive (IO). These cells express 5-HT(2)Rs on their soma and dendrites, and the IO receives extensive serotonergic input. We find that the excitatory synaptic inputs onto IO cells are strongly suppressed by serotonin receptor agonists as well as release of endogenous serotonin. Both 5-HT(2)Rs and 5-HT(1B)Rs contribute to this modulation by decreasing the probability of glutamate release from presynaptic boutons. The suppression by 5-HT(2)Rs is of particular interest because it is prevented by CB(1)R antagonists, and 5-HT(2)Rs are thought to be located only postsynaptically on IO cells. Our results indicate that serotonin activates 5-HT(2)Rs on IO neurons, thereby releasing endocannabinoids that act retrogradely to suppress glutamate release by activating presynaptic CB(1)Rs. These findings establish a link between serotonin signaling and endocannabinoid signaling. Based on the extensive distribution of 5-HT(2)Rs and CB(1)Rs, it seems likely that this mechanism could mediate many of the actions of 5-HT(2)Rs throughout the brain.
Figures
References
-
- Aghajanian GK, Marek GJ. Serotonin and hallucinogens. Neuropsychopharmacology. 1999;21:16S–23S. - PubMed
-
- Azizi SA, Woodward DJ. Inferior olivary nuclear complex of the rat: morphology and comments on the principles of organization within the olivocerebellar system. J Comp Neurol. 1987;263:467–484. - PubMed
-
- Barnes NM, Sharp T. A review of central 5-HT receptors and their function. Neuropharmacology. 1999;38:1083–1152. - PubMed
-
- Bishop GA, Ho RH. Cell bodies of origin of serotonin-immunoreactive afferents to the inferior olivary complex of the rat. Brain Res. 1986;399:369–373. - PubMed
-
- Bourrat F, Sotelo C. Postnatal development of the inferior olivary complex in the rat. I. An electron microscopic study of the medial accessory olive. Brain Res. 1983;284:291–310. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources