

The amphioxus genome illuminates vertebrate origins and cephalochordate biology
- PMID: 18562680
- PMCID: PMC2493399
- DOI: 10.1101/gr.073676.107
The amphioxus genome illuminates vertebrate origins and cephalochordate biology
Erratum in
- Genome Res. 2008 Aug;18(8):1380
Abstract
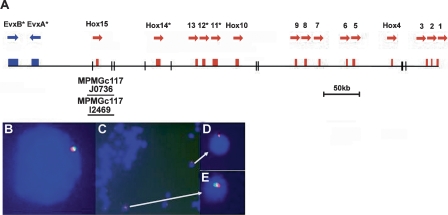
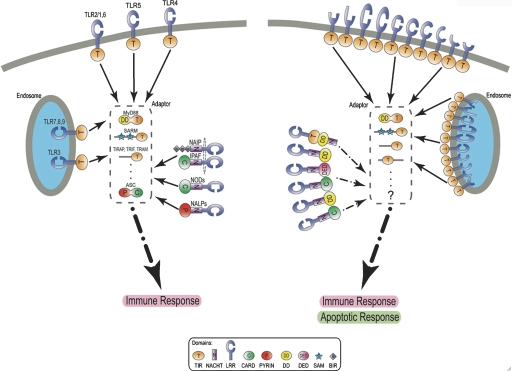
Cephalochordates, urochordates, and vertebrates evolved from a common ancestor over 520 million years ago. To improve our understanding of chordate evolution and the origin of vertebrates, we intensively searched for particular genes, gene families, and conserved noncoding elements in the sequenced genome of the cephalochordate Branchiostoma floridae, commonly called amphioxus or lancelets. Special attention was given to homeobox genes, opsin genes, genes involved in neural crest development, nuclear receptor genes, genes encoding components of the endocrine and immune systems, and conserved cis-regulatory enhancers. The amphioxus genome contains a basic set of chordate genes involved in development and cell signaling, including a fifteenth Hox gene. This set includes many genes that were co-opted in vertebrates for new roles in neural crest development and adaptive immunity. However, where amphioxus has a single gene, vertebrates often have two, three, or four paralogs derived from two whole-genome duplication events. In addition, several transcriptional enhancers are conserved between amphioxus and vertebrates--a very wide phylogenetic distance. In contrast, urochordate genomes have lost many genes, including a diversity of homeobox families and genes involved in steroid hormone function. The amphioxus genome also exhibits derived features, including duplications of opsins and genes proposed to function in innate immunity and endocrine systems. Our results indicate that the amphioxus genome is elemental to an understanding of the biology and evolution of nonchordate deuterostomes, invertebrate chordates, and vertebrates.
Figures




References
-
- Adams M.D., Celniker S.E., Holt R.A., Evans C.A., Gocayne J.D., Amanatides P.G., Scherer S.E., Li P.W., Hoskins R.A., Galle R.F., et al. The genome sequence of Drosophila melanogaster. Science. 2000;287:2185–2195. - PubMed
-
- Aparicio S., Chapman J., Stupka E., Putnam N., Chia J.-m., Dehal P., Christoffels A., Rash S., Hoon S., Smit A., et al. Whole-genome shotgun assembly and analysis of the genome of Fugu rubripes. Science. 2002;297:1301–1310. - PubMed
-
- Arendt D., Tessmar-Raible K., Snyman H., Dorresteijn A.W., Wittbrodt J. Ciliary photoreceptors with a vertebrate-type opsin in an invertebrate brain. Science. 2004;306:869–871. - PubMed
-
- Beaster-Jones L., Schubert M., Holland L.Z. Cis-regulation of the amphioxus engrailed gene: Insights into evolution of a muscle-specific enhancer. Mech. Dev. 2007;124:532–542. - PubMed
-
- Boraschi D., Tagliabue A. The interleukin-1 receptor family. Vitam. Horm. 2006;74:229–254. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials