

Manganese transport in eukaryotes: the role of DMT1
- PMID: 18565586
- PMCID: PMC2501114
- DOI: 10.1016/j.neuro.2008.04.022
Manganese transport in eukaryotes: the role of DMT1
Abstract
Manganese (Mn) is a transition metal that is essential for normal cell growth and development, but is toxic at high concentrations. While Mn deficiency is uncommon in humans, Mn toxicity is known to be readily prevalent due to occupational overexposure in miners, smelters and possibly welders. Excessive exposure to Mn can cause Parkinson's disease-like syndrome; patients typically exhibit extrapyramidal symptoms that include tremor, rigidity and hypokinesia [Calne DB, Chu NS, Huang CC, Lu CS, Olanow W. Manganism and idiopathic parkinsonism: similarities and differences. Neurology 1994;44(9):1583-6; Dobson AW, Erikson KM, Aschner M. Manganese neurotoxicity. Ann NY Acad Sci 2004;1012:115-28]. Mn-induced motor neuron diseases have been the subjects of numerous studies; however, this review is not intended to discuss its neurotoxic potential or its role in the etiology of motor neuron disorders. Rather, it will focus on Mn uptake and transport via the orthologues of the divalent metal transporter (DMT1) and its possible implications to Mn toxicity in various categories of eukaryotic systems, such as in vitro cell lines, in vivo rodents, the fruitfly, Drosophila melanogaster, the honeybee, Apis mellifera L., the nematode, Caenorhabditis elegans and the baker's yeast, Saccharomyces cerevisiae.
Figures
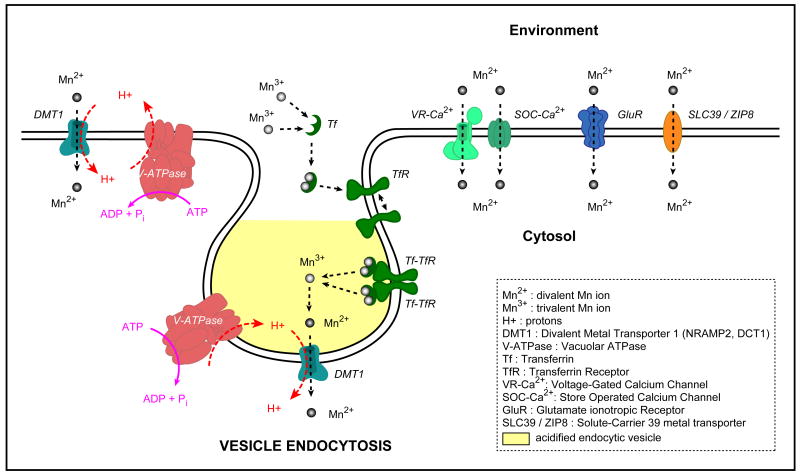

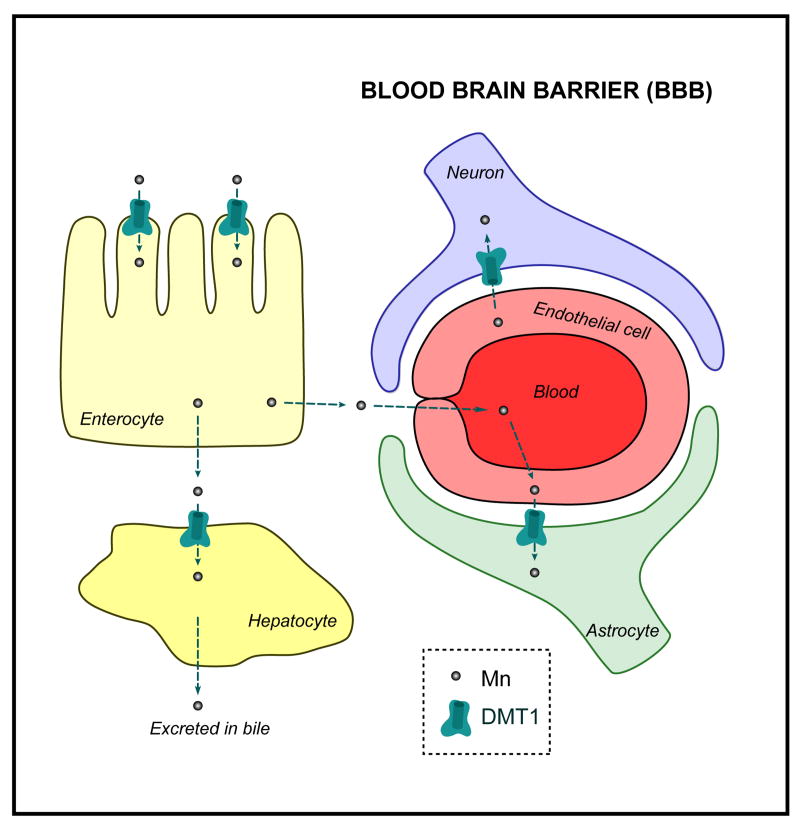
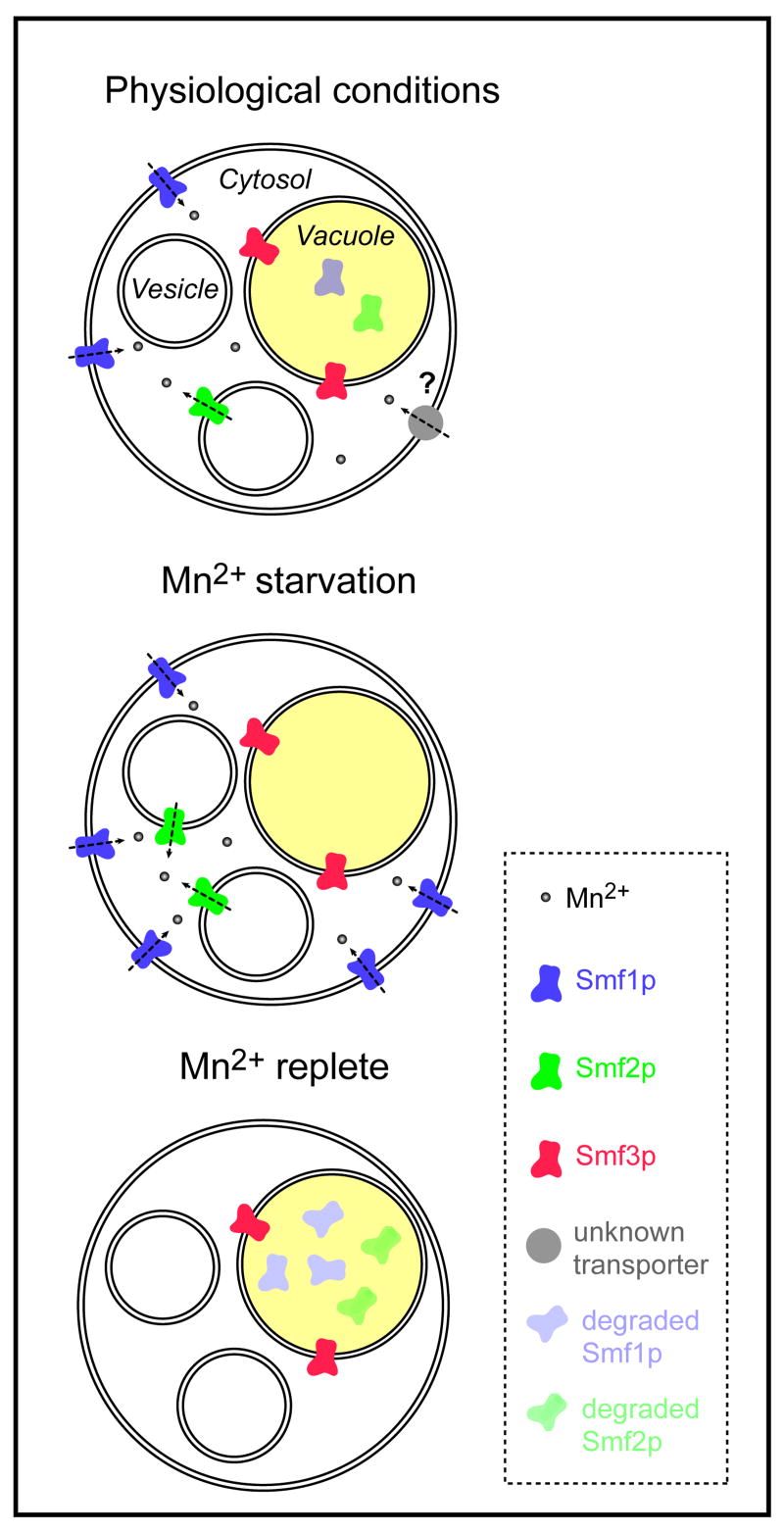
References
-
- Addess KJ, Basilion JP, Klausner RD, Rouault TA, Pardi A. Structure and dynamics of the iron responsive element RNA: implications for binding of the RNA by iron regulatory binding proteins. J Mol Biol. 1997;274(1):72–83. - PubMed
-
- Andrews NC. The iron transporter DMT1. Int J Biochem Cell Biol. 1999;31(10):991–4. - PubMed
-
- Aschner M, Aschner JL. Manganese neurotoxicity: cellular effects and blood-brain barrier transport. Neurosci Biobehav Rev. 1991;15(3):333–40. - PubMed
-
- Aschner M, Gannon M. Manganese (Mn) transport across the rat blood-brain barrier: saturable and transferrin-dependent transport mechanisms. Brain Res Bull. 1994;33(3):345–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
