

Lin-28 interaction with the Let-7 precursor loop mediates regulated microRNA processing
- PMID: 18566191
- PMCID: PMC2491462
- DOI: 10.1261/rna.1155108
Lin-28 interaction with the Let-7 precursor loop mediates regulated microRNA processing
Abstract
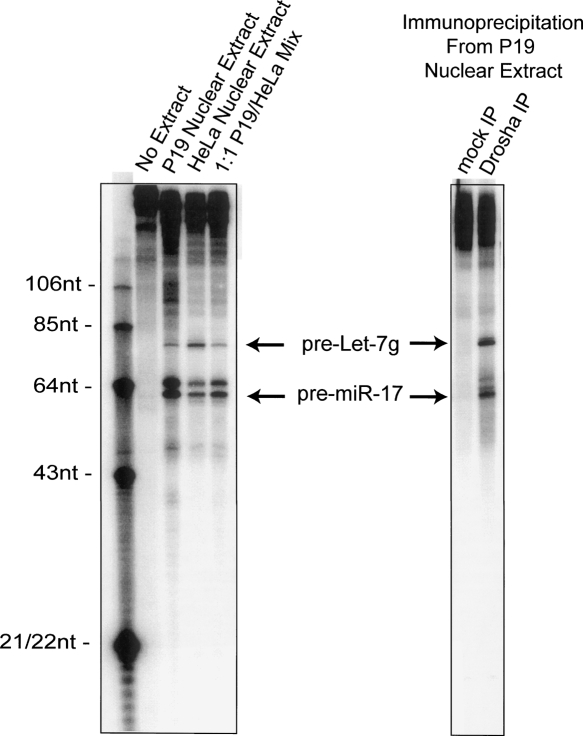
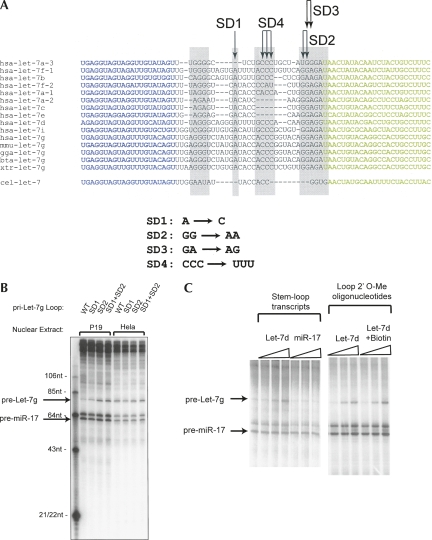
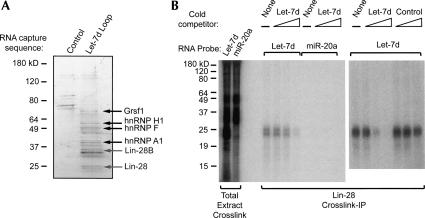
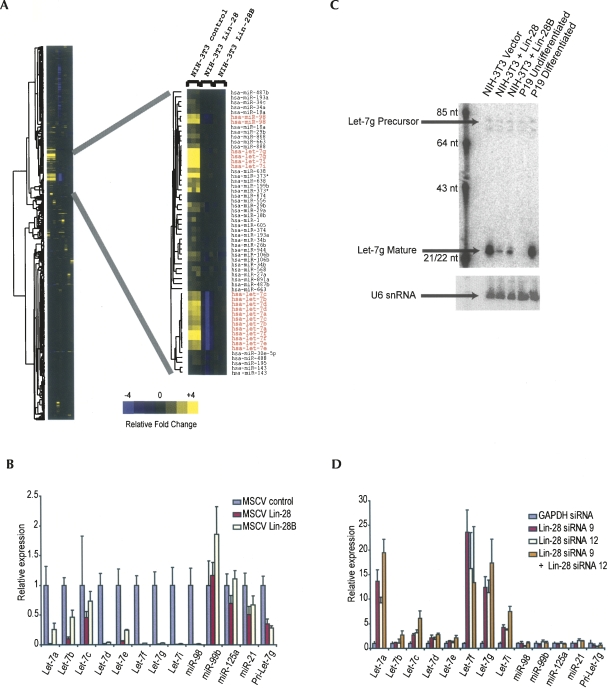
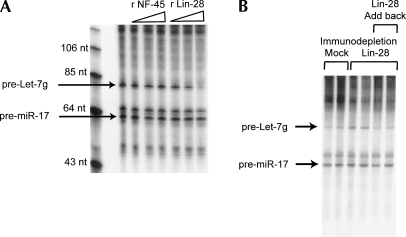
A hallmark of mammalian embryonic development is the widespread induction of microRNA (miRNA) expression. Surprisingly, the transcription of many of these small, noncoding RNAs is unchanged through development; rather, a post-transcriptional regulatory event prevents accumulation of the mature miRNA species. Here, we present a biochemical framework for the regulated production of the Let-7 family of miRNAs. Embryonic cells contain a Drosha Inhibitor that prevents processing of the Let-7 primary transcript. This inhibitor specifically binds to conserved nucleotides in the loop region of the Let-7 precursor, and competitor RNAs that mimic the binding site restore Let-7 processing. We have identified the Drosha Inhibitor as the embryonic stem cell specific protein Lin-28. Lin-28 has been previously implicated in developmental regulatory pathways in Caenorhabditis elegans, and it promotes reprogramming of human somatic cells into pluripotent stem cells. Our findings outline a microRNA post-transcriptional regulatory network and establish a novel role for the miRNA precursor loop in the regulated production of mature Let-7.
Figures
References
-
- Ambros V. A hierarchy of regulatory genes controls a larva-to-adult developmental switch in C. elegans . Cell. 1989;57:49–57. - PubMed
-
- Ambros V., Horvitz H.R. Heterochronic mutants of the nematode Caenorhabditis elegans . Science. 1984;226:409–416. - PubMed
-
- Balzer E., Moss E.G. Localization of the developmental timing regulator Lin28 to mRNP complexes, P-bodies and stress granules. RNA Biol. 2007;4:16–25. - PubMed
-
- Bartel D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell. 2004;116:281–297. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials