

Revisiting the GroEL-GroES reaction cycle via the symmetric intermediate implied by novel aspects of the GroEL(D398A) mutant
- PMID: 18567584
- PMCID: PMC3259786
- DOI: 10.1074/jbc.M802542200
Revisiting the GroEL-GroES reaction cycle via the symmetric intermediate implied by novel aspects of the GroEL(D398A) mutant
Abstract
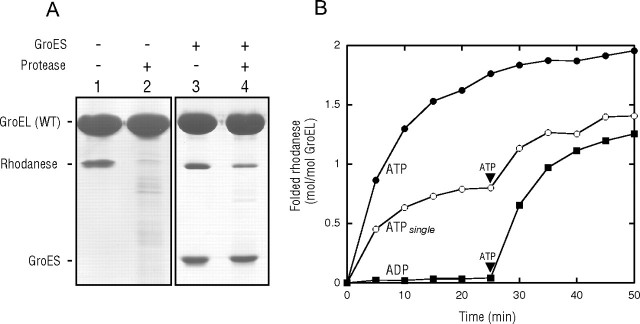
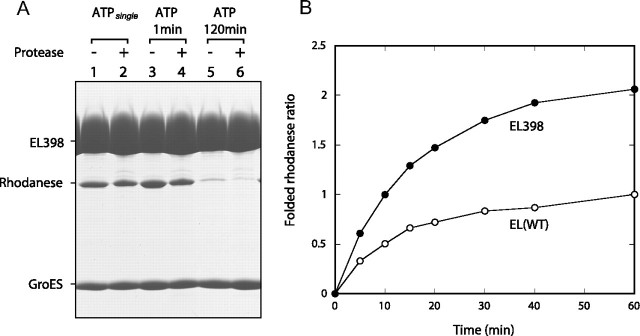
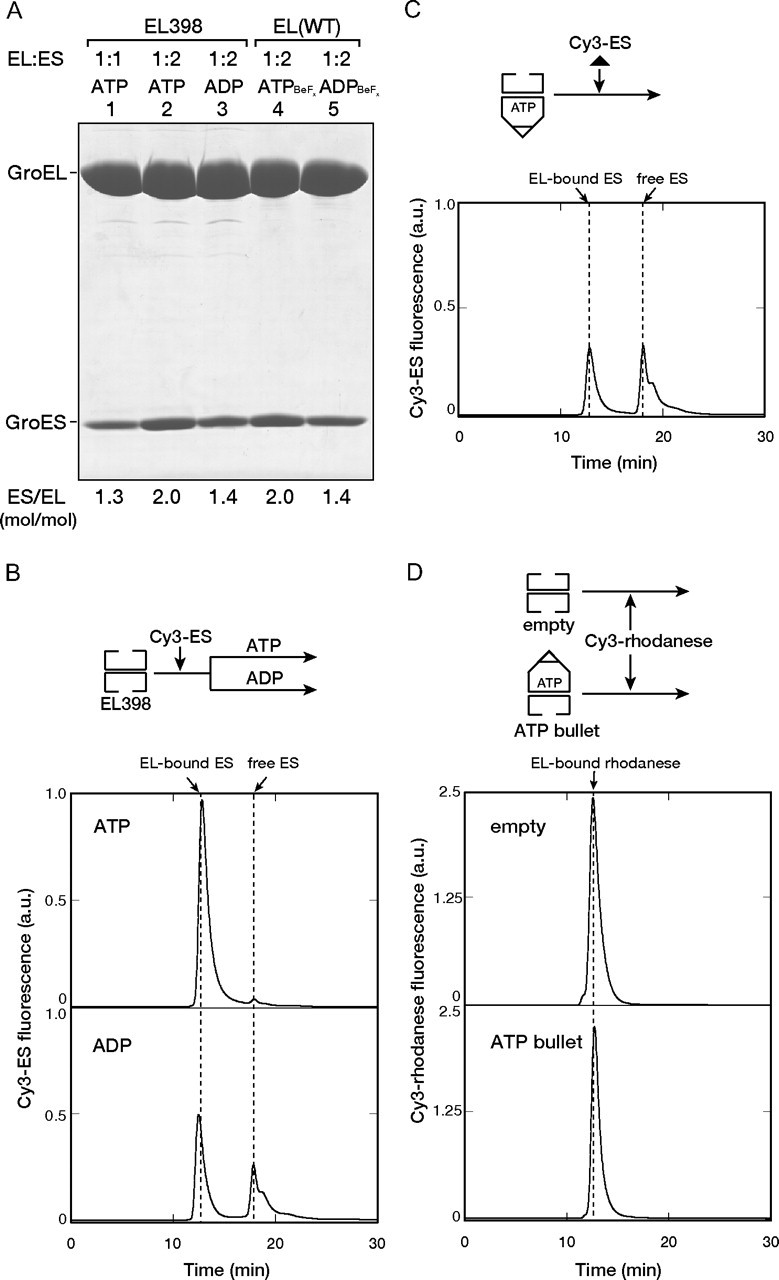
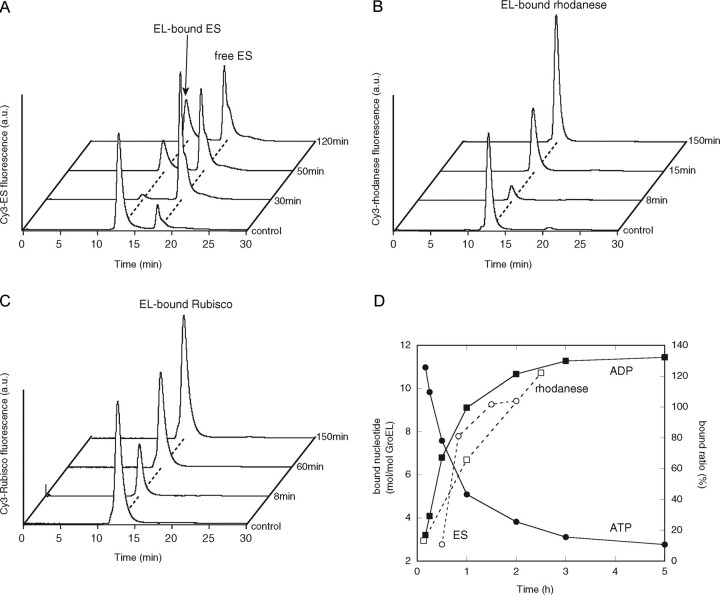
The Escherichia coli chaperonin GroEL is a double-ring chaperone that assists in protein folding with the aid of GroES and ATP. It is believed that GroEL alternates the folding-active rings and that the substrate protein (and GroES) can bind to the open trans-ring only after ATP in the cis-ring is hydrolyzed. However, we found that a substrate protein prebound to the trans-ring remained bound during the first ATP cycle, and this substrate was assisted by GroEL-GroES when the second cycle began. Moreover, a slow ATP-hydrolyzing GroEL mutant (D398A) in the ATP-bound form bound a substrate protein and GroES to the trans-ring. The apparent discrepancy with the results from an earlier study (Rye, H. S., Roseman, A. M., Chen, S., Furtak, K., Fenton, W. A., Saibil, H. R., and Horwich, A. L. (1999) Cell 97, 325-338) can be explained by the previously unnoticed fact that the ATP-bound form of the D398A mutant exists as a symmetric 1:2 GroEL-GroES complex (the "football"-shaped complex) and that the substrate protein (and GroES) in the medium is incorporated into the complex only after the slow turnover. In light of these results, the current model of the GroEL-GroES reaction cycle via the asymmetric 1:1 GroEL-GroES complex deserves reexamination.
Figures
References
-
- Hartl, F. U., and Hayer-Hartl, M. (2002) Science 295 1852–1858 - PubMed
-
- Young, J. C., Agashe, V. R., Siegers, K., and Hartl, F. U. (2004) Nat. Rev. Mol. Cell Biol. 5 781–791 - PubMed
-
- Thirumalai, D., and Lorimer, G. H. (2001) Annu. Rev. Biophys. Biomol. Struct. 30 245–269 - PubMed
-
- Taguchi, H. (2005) J. Biochem. (Tokyo) 137 543–549 - PubMed
-
- Horwich, A. L., Fenton, W. A., Chapman, E., and Farr, G. W. (2007) Annu. Rev. Cell Dev. Biol. 23 115–145 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
