

Protonation and sugar binding to LacY
- PMID: 18567672
- PMCID: PMC3021437
- DOI: 10.1073/pnas.0803577105
Protonation and sugar binding to LacY
Abstract
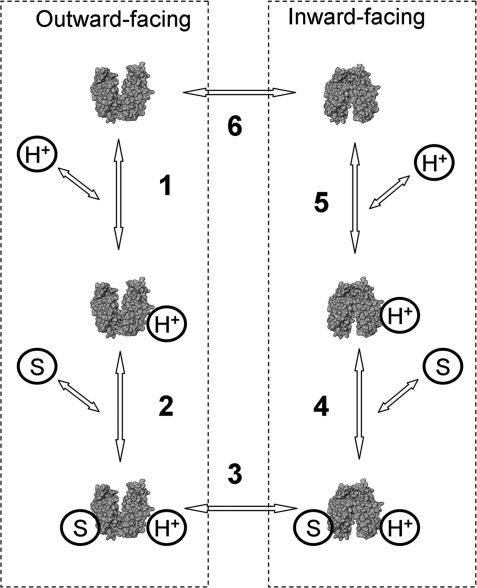
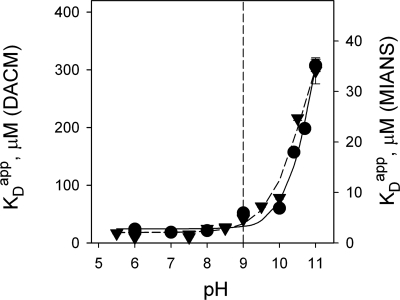
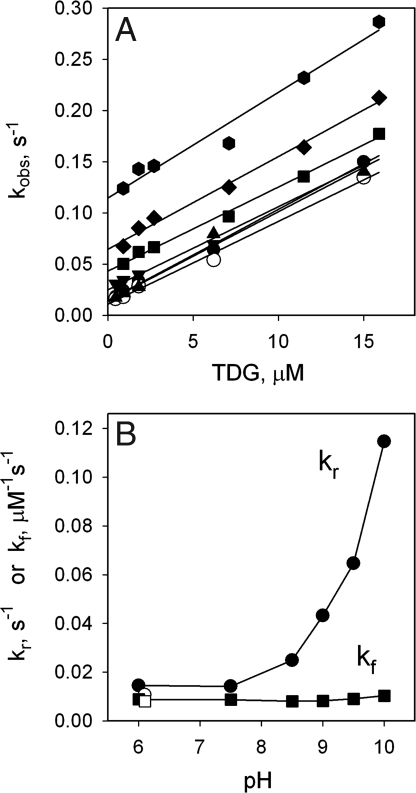
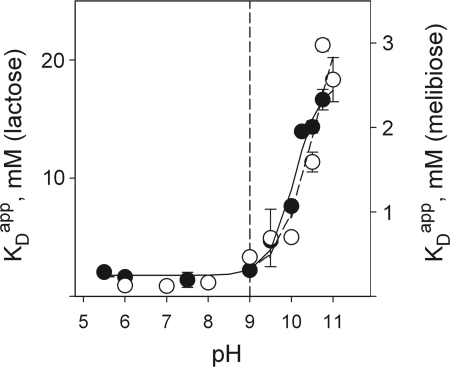
The effect of bulk-phase pH on the apparent affinity (K(d)(app)) of purified wild-type lactose permease (LacY) for sugars was studied. K(d)(app) values were determined by ligand-induced changes in the fluorescence of either of two covalently bound fluorescent reporters positioned away from the sugar-binding site. K(d)(app) for three different galactopyranosides was determined over a pH range from 5.5 to 11. A remarkably high pK(a) of approximately 10.5 was obtained for all sugars. Kinetic data for thiodigalactoside binding measured from pH 6 to 10 show that decreased affinity for sugar at alkaline pH is due specifically to increased reverse rate. A similar effect was also observed with nitrophenylgalactoside by using a direct binding assay. Because affinity for sugar remains constant from pH 5.5 to pH 9.0, it follows that LacY is fully protonated with respect to sugar binding under physiological conditions of pH. The results are consistent with the conclusion that LacY is protonated before sugar binding during lactose/H(+) symport in either direction across the membrane.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
-
- Saier MH, Jr, et al. The major facilitator superfamily. J Mol Microbiol Biotechnol. 1999;1:257–279. - PubMed
-
- Saier MH., Jr Families of transmembrane sugar transport proteins. Mol Microbiol. 2000;35:699–710. - PubMed
-
- West IC. Lactose transport coupled to proton movements in Escherichia coli. Biochem Biophys Res Commun. 1970;41:655–661. - PubMed
-
- Kaback HR, Sahin-Toth M, Weinglass AB. The kamikaze approach to membrane transport. Nat Rev Mol Cell Biol. 2001;2:610–620. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
