

Endothelial-specific ablation of serum response factor causes hemorrhaging, yolk sac vascular failure, and embryonic lethality
- PMID: 18570667
- PMCID: PMC2442838
- DOI: 10.1186/1471-213X-8-65
Endothelial-specific ablation of serum response factor causes hemorrhaging, yolk sac vascular failure, and embryonic lethality
Abstract
Background: Serum response factor (SRF), a member of the MADS box family of nuclear transcription factors, plays an important role in cardiovascular development and function. Numerous studies demonstrate a central role for SRF in regulating smooth and cardiac muscle cell gene expression. Consistent with this, loss of SRF function blocks differentiation of coronary vascular smooth muscle cells from proepicardial precursors, indicating SRF is necessary for coronary vasculogenesis. The role of SRF in endothelial cell contribution during early vascular development, however, has not been addressed. To investigate this, we generated transgenic mice lacking expression of SRF in endothelial cells. Mice expressing Cre recombinase (Tie2Cre+) under Tie2 promoter control were bred to mice homozygous for Srf alleles containing loxP recombination sites within the Srf gene (Srff/f). Tie2 is a tyrosine kinase receptor expressed predominantly on endothelial cells that mediates signalling during different stages of blood vessel remodelling. Resulting embryos were harvested at specific ages for observation of physical condition and analysis of genotype.
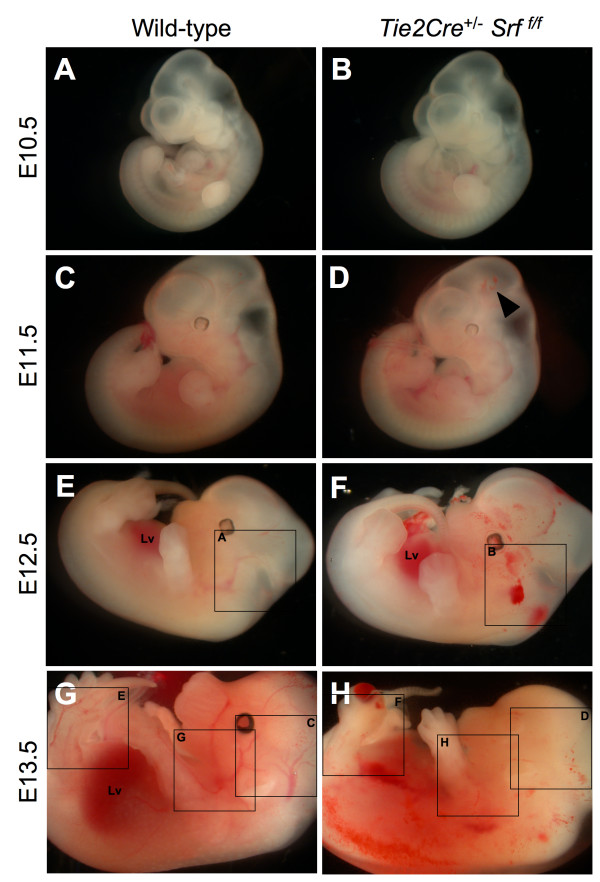
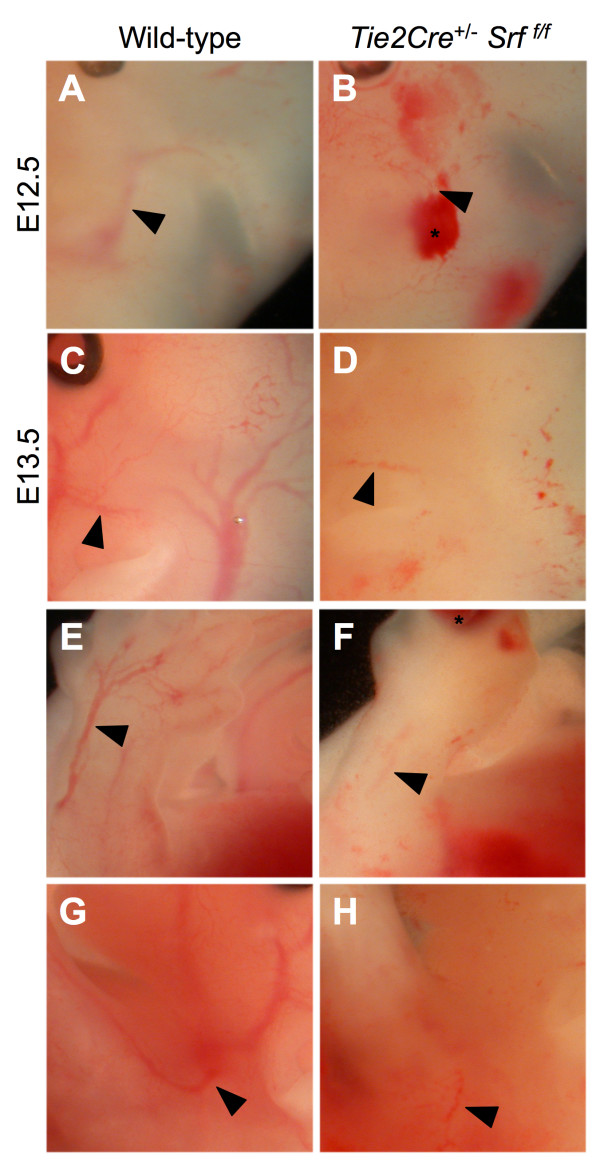
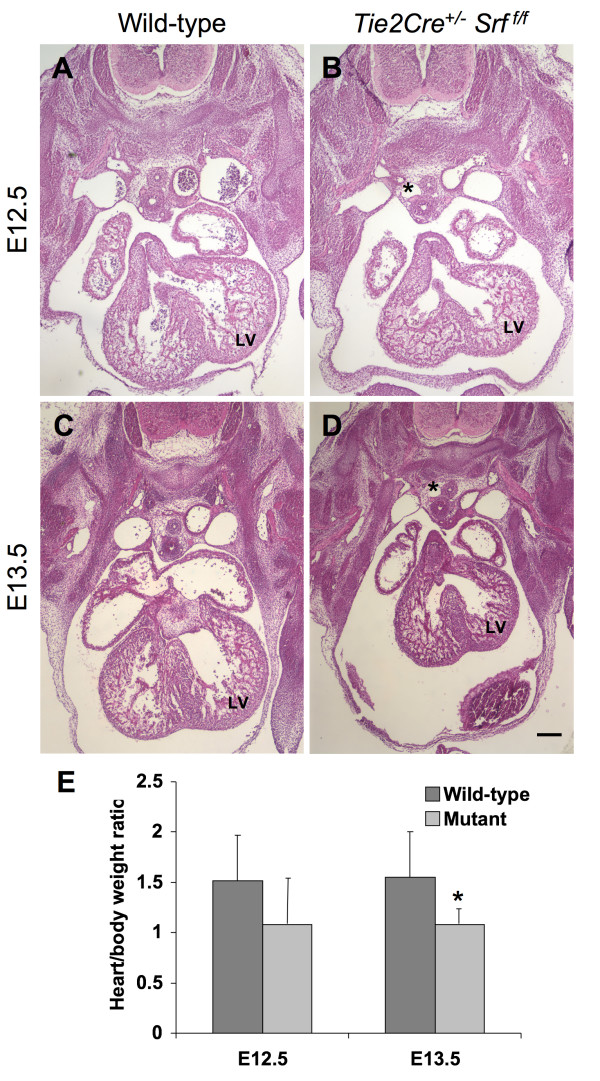
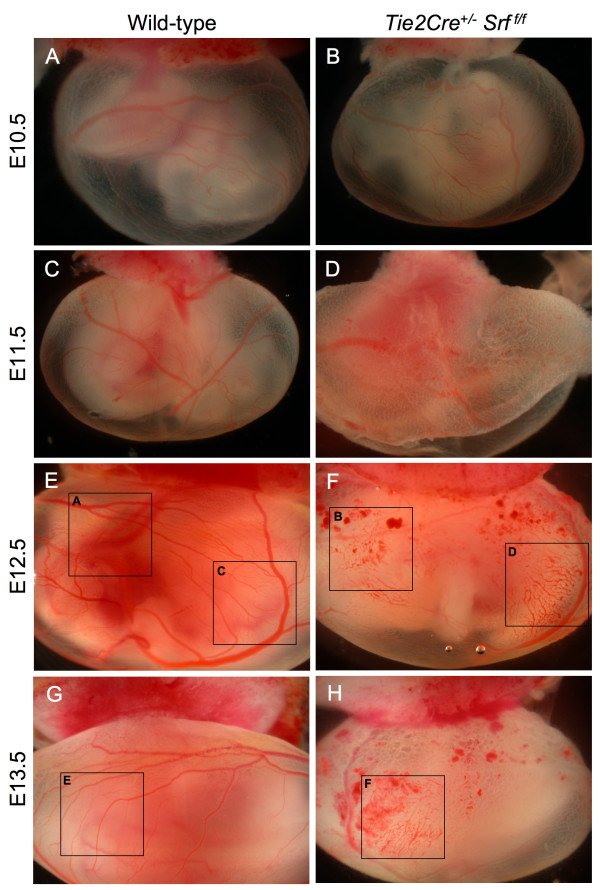
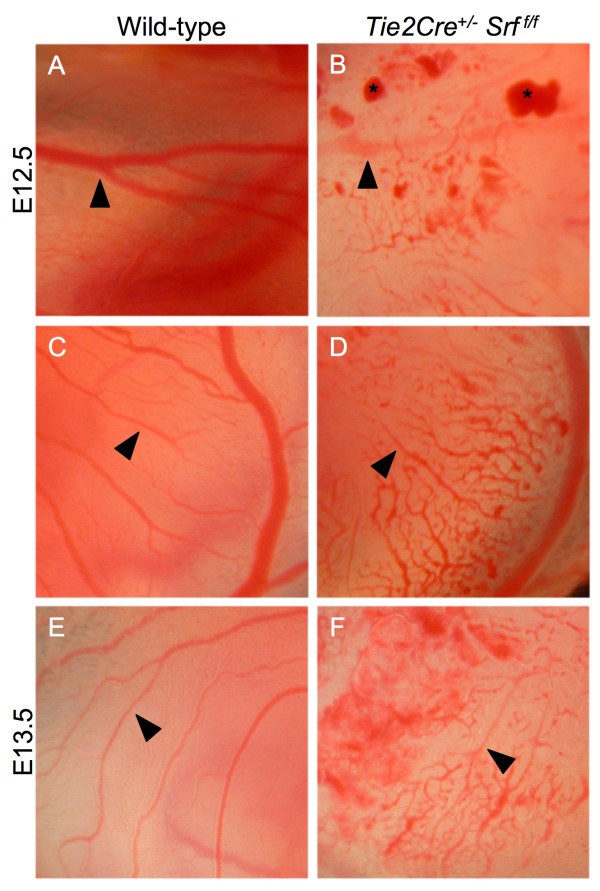
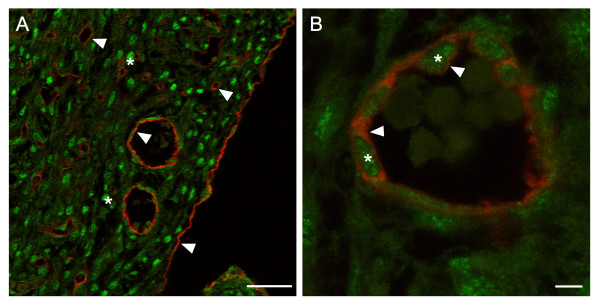
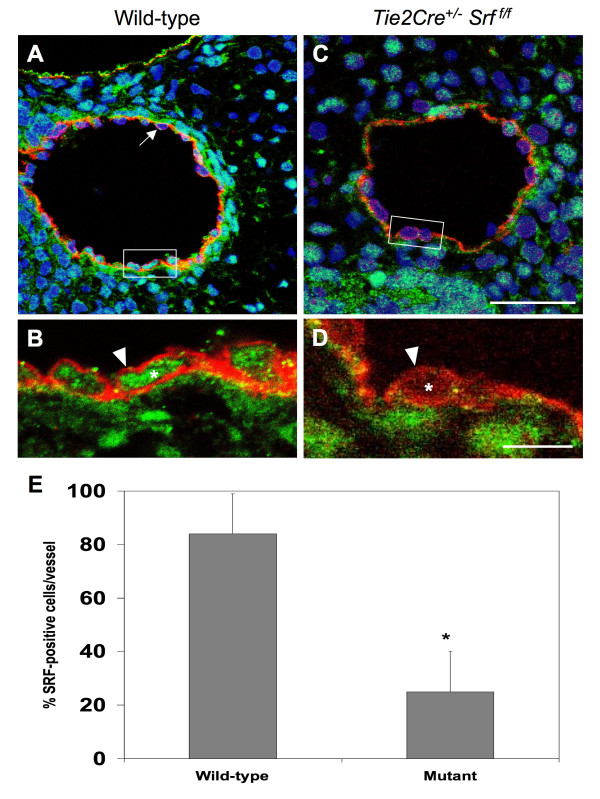
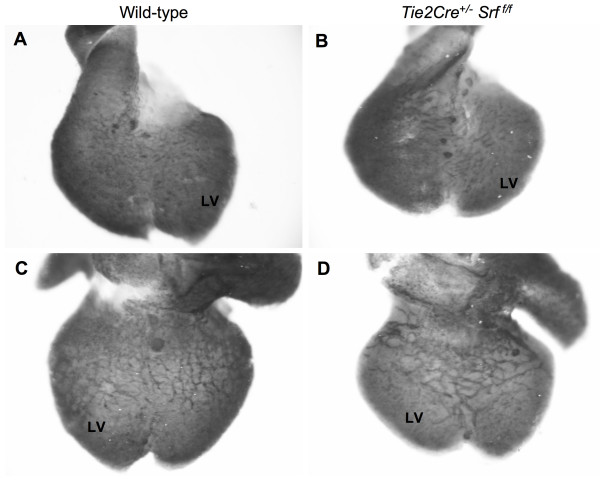
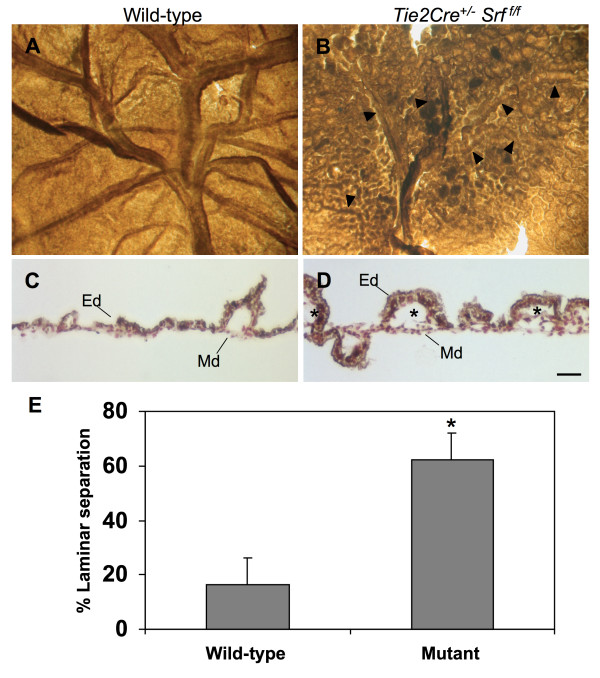
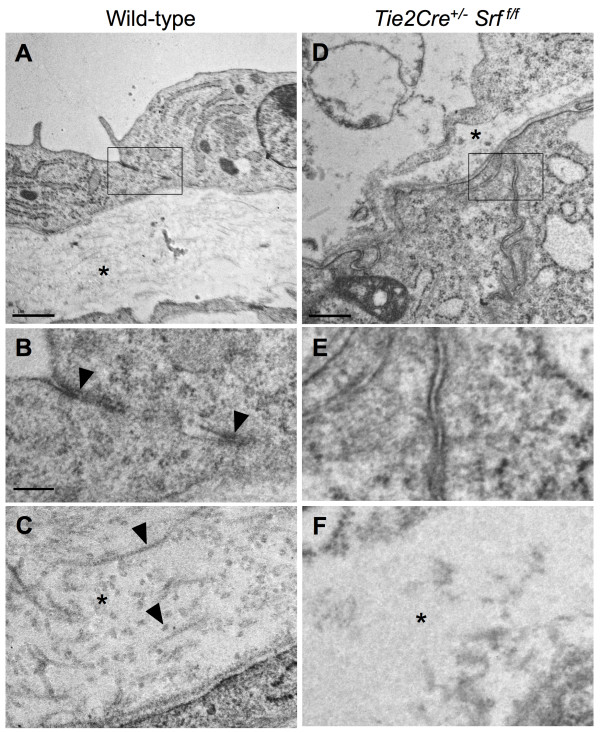
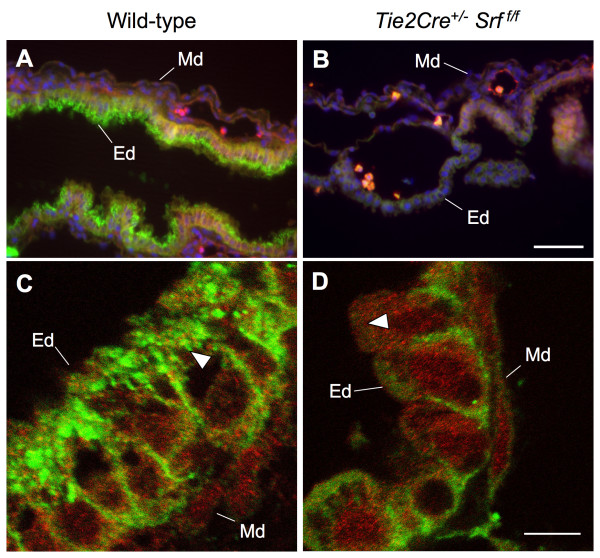
Results: Tie2Cre+/-Srff/f embryos appeared to develop normally compared to wild-type littermates until embryonic day 10.5 (E10.5). Beginning at E11.5, Tie2Cre+/-Srff/f embryos exhibited cerebrovascular hemorrhaging and severely disrupted vascular networks within the yolk sac. Hemorrhaging in mutant embryos became more generalized with age, and by E14.5, most Tie2Cre+/-Srff/f embryos observed were nonviable and grossly necrotic. Hearts of mutant embryos were smaller relative to overall body weight compared to wild-type littermates. Immunohistochemical analysis revealed the presence of vascular endothelial cells; however, vessels failed to undergo appropriate remodelling. Initial analysis by electron microscopy suggested a lack of appropriate cell-cell contacts between endothelial cells. Consistent with this, disrupted E-cadherin staining patterns were observed in mutant embryos.
Conclusion: These results provide the first in vivo evidence in support of a role for SRF in endothelial cell function and strongly suggest SRF is required for appropriate vascular remodelling.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
