

Nuclear trapping shapes the terminal gradient in the Drosophila embryo
- PMID: 18571412
- PMCID: PMC2500156
- DOI: 10.1016/j.cub.2008.05.034
Nuclear trapping shapes the terminal gradient in the Drosophila embryo
Abstract
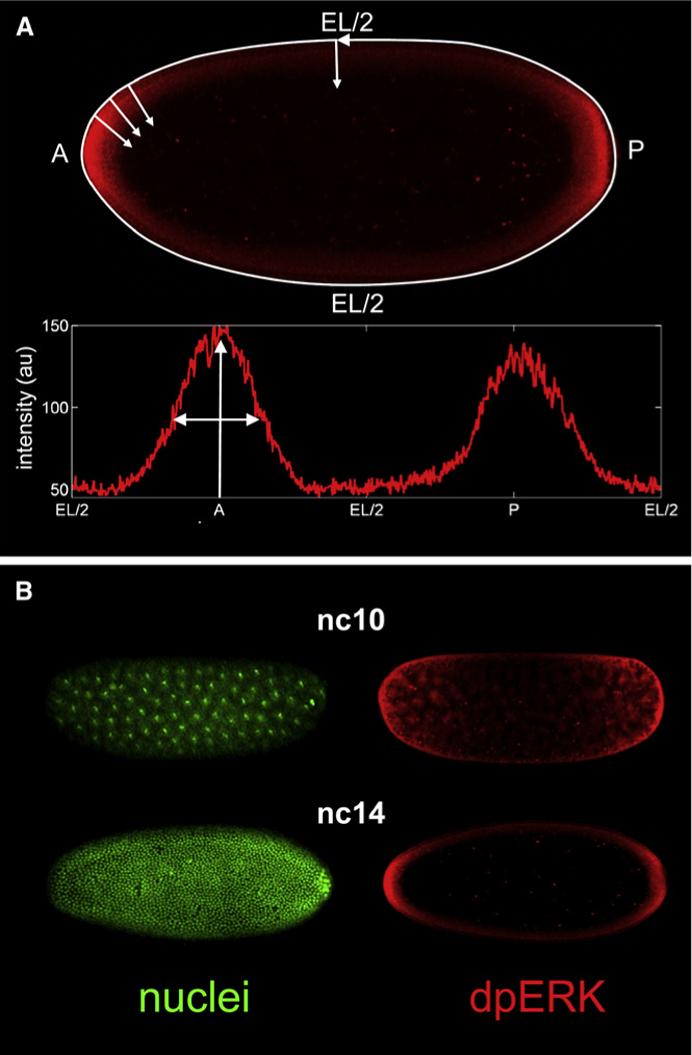
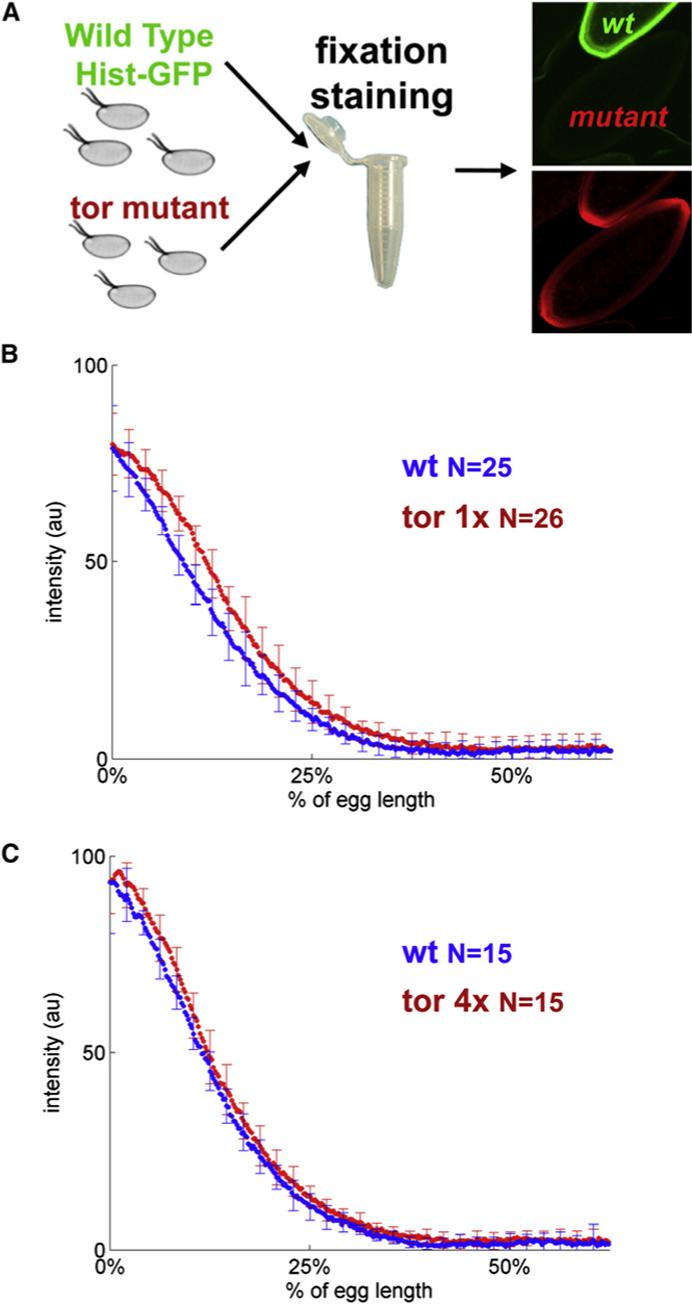
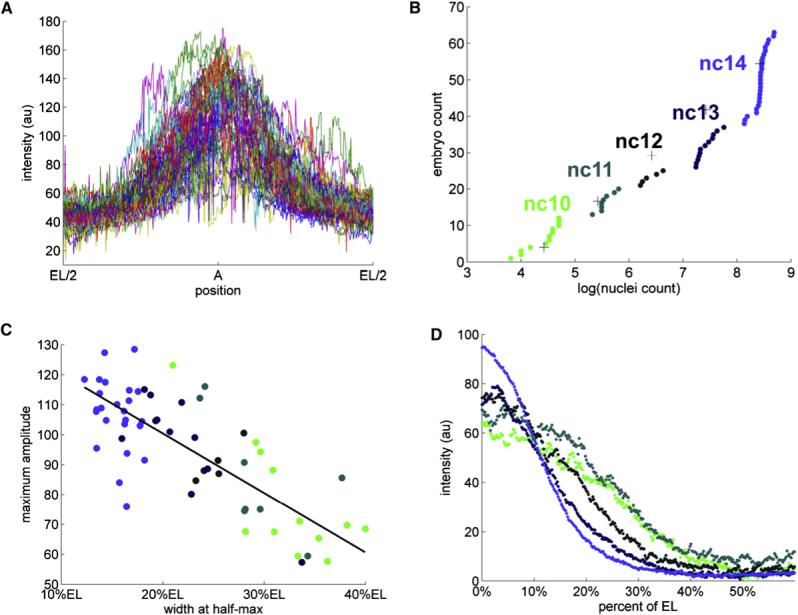
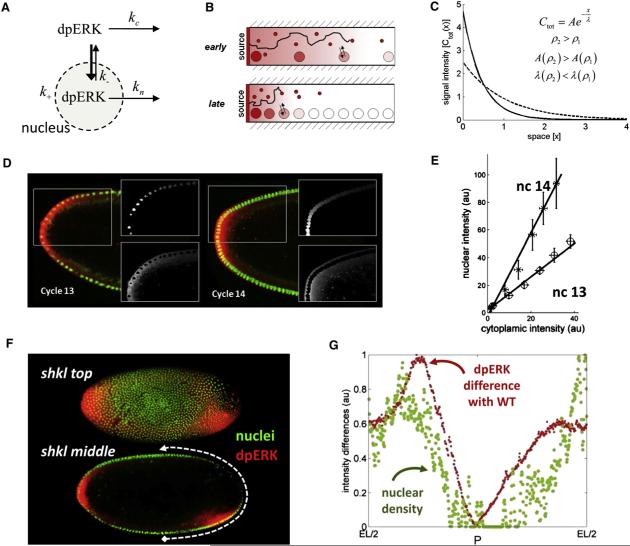
Patterning of the terminal regions of the Drosophila embryo relies on the gradient of phosphorylated ERK/MAPK (dpERK), which is controlled by the localized activation of the Torso receptor tyrosine kinase [1-4]. This model is supported by a large amount of data, but the gradient itself has never been quantified. We present the first measurements of the dpERK gradient and establish a new intracellular layer of its regulation. Based on the quantitative analysis of the spatial pattern of dpERK in mutants with different levels of Torso as well as the dynamics of the wild-type dpERK pattern, we propose that the terminal-patterning gradient is controlled by a cascade of diffusion-trapping modules. A ligand-trapping mechanism establishes a sharply localized pattern of the Torso receptor occupancy on the surface of the embryo. Inside the syncytial embryo, nuclei play the role of traps that localize diffusible dpERK. We argue that the length scale of the terminal-patterning gradient is determined mainly by the intracellular module.
Figures
Similar articles
-
The torso receptor localizes as well as transduces the spatial signal specifying terminal body pattern in Drosophila.Nature. 1993 Mar 11;362(6416):152-5. doi: 10.1038/362152a0. Nature. 1993. PMID: 8450886
-
Trunk cleavage is essential for Drosophila terminal patterning and can occur independently of Torso-like.Nat Commun. 2014 Mar 3;5:3419. doi: 10.1038/ncomms4419. Nat Commun. 2014. PMID: 24584029
-
A MAPK docking site is critical for downregulation of Capicua by Torso and EGFR RTK signaling.EMBO J. 2007 Feb 7;26(3):668-77. doi: 10.1038/sj.emboj.7601532. Epub 2007 Jan 25. EMBO J. 2007. PMID: 17255944 Free PMC article.
-
Pattern formation by receptor tyrosine kinases: analysis of the Gurken gradient in Drosophila oogenesis.Curr Opin Genet Dev. 2011 Dec;21(6):719-25. doi: 10.1016/j.gde.2011.07.009. Epub 2011 Aug 19. Curr Opin Genet Dev. 2011. PMID: 21862318 Free PMC article. Review.
-
Functions and mechanisms of receptor tyrosine kinase Torso signaling: lessons from Drosophila embryonic terminal development.Dev Dyn. 2005 Mar;232(3):656-72. doi: 10.1002/dvdy.20295. Dev Dyn. 2005. PMID: 15704136 Free PMC article. Review.
Cited by
-
β-arrestin Kurtz inhibits MAPK and Toll signalling in Drosophila development.EMBO J. 2010 Oct 6;29(19):3222-35. doi: 10.1038/emboj.2010.202. Epub 2010 Aug 27. EMBO J. 2010. PMID: 20802461 Free PMC article.
-
Rapid Dynamics of Signal-Dependent Transcriptional Repression by Capicua.Dev Cell. 2020 Mar 23;52(6):794-801.e4. doi: 10.1016/j.devcel.2020.02.004. Epub 2020 Mar 5. Dev Cell. 2020. PMID: 32142631 Free PMC article.
-
Divergent effects of intrinsically active MEK variants on developmental Ras signaling.Nat Genet. 2017 Mar;49(3):465-469. doi: 10.1038/ng.3780. Epub 2017 Feb 6. Nat Genet. 2017. PMID: 28166211 Free PMC article.
-
Dynamics of an incoherent feedforward loop drive ERK-dependent pattern formation in the early Drosophila embryo.Development. 2023 Sep 1;150(17):dev201818. doi: 10.1242/dev.201818. Epub 2023 Sep 1. Development. 2023. PMID: 37602510 Free PMC article.
-
The role of regulated mRNA stability in establishing bicoid morphogen gradient in Drosophila embryonic development.PLoS One. 2011;6(9):e24896. doi: 10.1371/journal.pone.0024896. Epub 2011 Sep 16. PLoS One. 2011. PMID: 21949782 Free PMC article.
References
-
- Casanova J, Struhl G. The torso receptor localizes as well as transduces the spatial signal specifying terminal body pattern in Drosophila. Nature. 1993;362:152–155. - PubMed
-
- Casanova J, Struhl G. Localized surface-activity of Torso, a receptor tyrosine kinase, specifies terminal body pattern in Drosophila. Genes Dev. 1989;3:2025–2038. - PubMed
-
- Sprenger F, Nusslein-Volhard C. Torso receptor activity is regulated by a diffusible ligand produced at the extracellular terminal regions of the Drosophila egg. Cell. 1992;71:987–1001. - PubMed
-
- Furriols M, Sprenger F, Casanova J. Variation in the number of activated torso receptors correlates with differential gene expression. Development. 1996;122:2313–2317. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
