

Review
doi: 10.1146/annurev.biophys.37.032807.125859.
PIP2 is a necessary cofactor for ion channel function: how and why?
Affiliations
- PMID: 18573078
- PMCID: PMC2692585
- DOI: 10.1146/annurev.biophys.37.032807.125859
Item in Clipboard
Review
PIP2 is a necessary cofactor for ion channel function: how and why?
Annu Rev Biophys.
2008.
Abstract
Phosphatidylinositol 4,5-bisphosphate (PIP2) is a minority phospholipid of the inner leaflet of plasma membranes. Many plasma membrane ion channels and ion transporters require PIP2 to function and can be turned off by signaling pathways that deplete PIP2. This review discusses the dependence of ion channels on phosphoinositides and considers possible mechanisms by which PIP2 and analogues regulate ion channel activity.
Figures
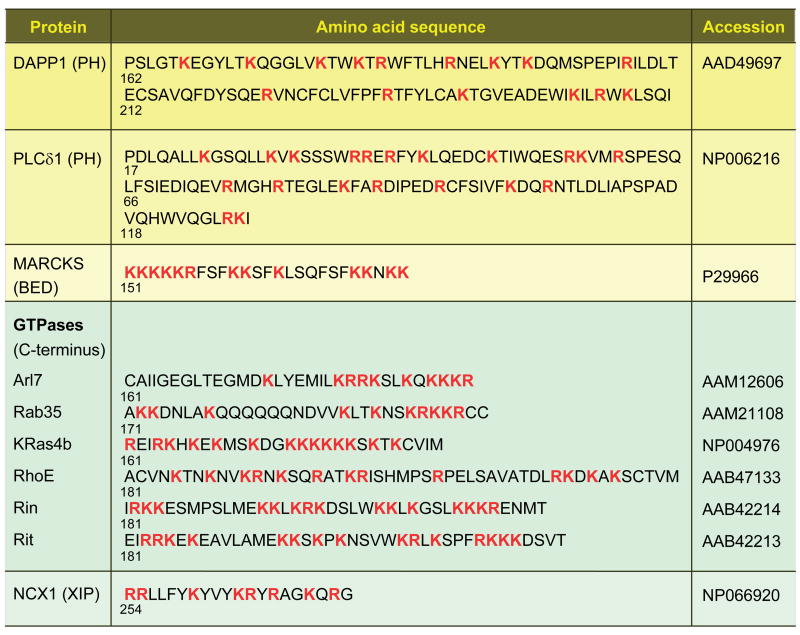
Crystal structure of the PH domain of DAPP1 (residues 162–261) bound to I(1,3,4,5)P4. a, The protein component alone, with the main chain drawn as a ribbon and five basic residues drawn in stick form. Their contacting nitrogens are drawn as CPK balls. b, The full complex with I(1,3,4,5)P4 represented in space-filling CPK form. The 1, 3, 4, and 5 phosphates are labeled. Colors: Blue nitrogen, red oxygen, orange phosphorus. Coordinates from (19).
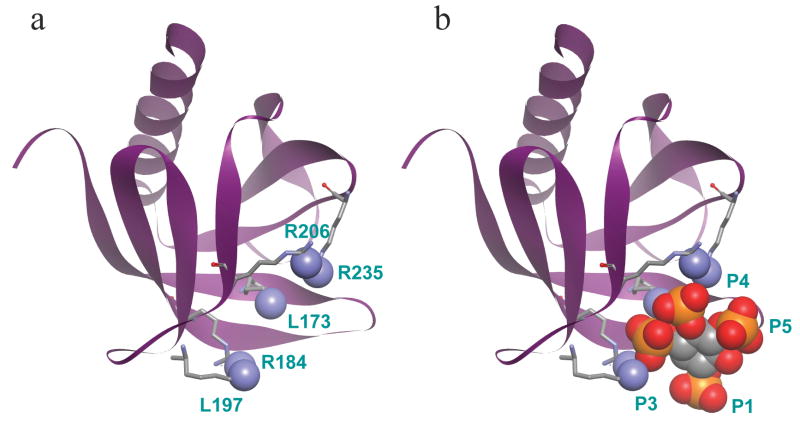
Amino acid sequences of some putative phosphoinositide-binding domains. All basic residues are bold and red. Shown are the PH domains of DAPP1 and PLCδ1, the BED domain of MARCKS; the C-terminus of several GTPases, and the XIP domain of the Na+-Ca2+ exchanger (NCX1). Numbers below each sequence indicate the initial residue number.
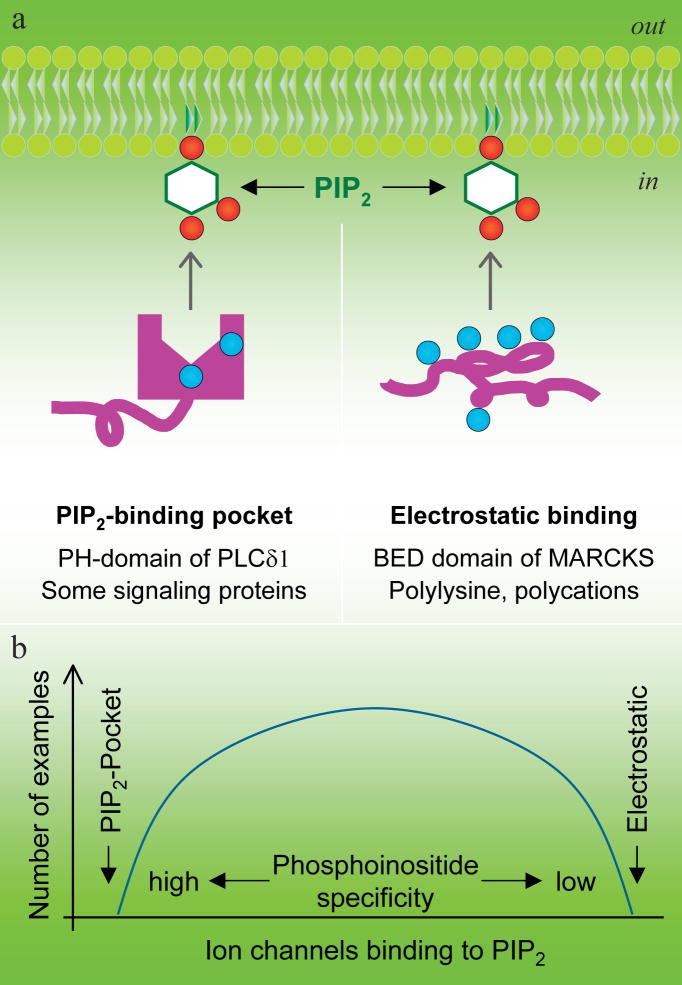
Two extreme models for selective versus diffuse binding of poly-phosphoinositides. a, Specific PIP2 interaction in a structured PIP2 binding pocket structure and nonselective electrostatic PIP2 attraction by polybasic peptides or polycations. b, A hypothesis suggesting a correlation between strong PIP2 binding and high PIP2 selectivity. The curve representing the number of channels in each category is completely hypothetical.
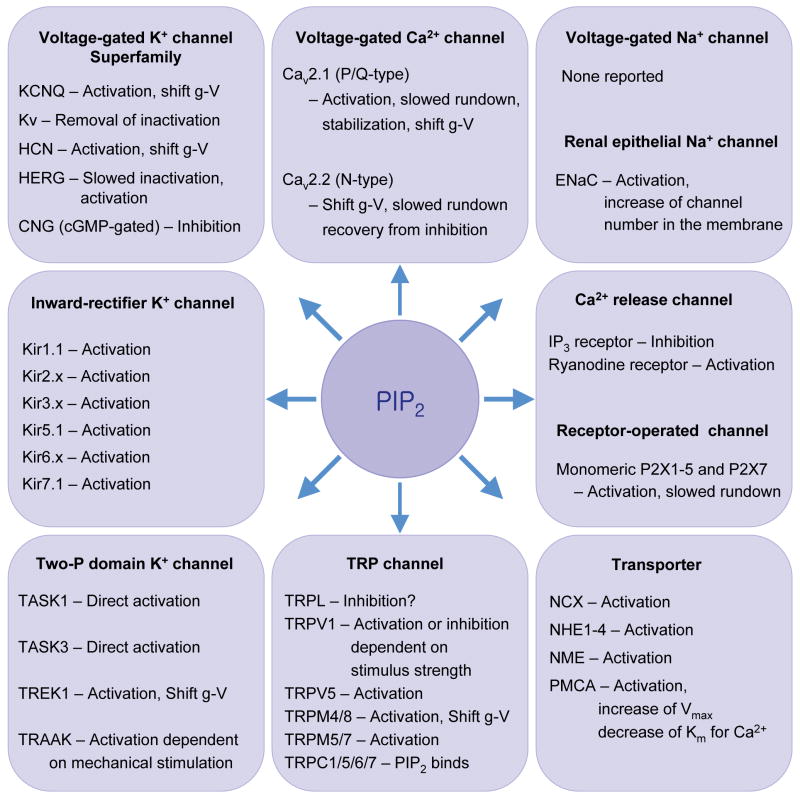
Ion channels and transporters sensitive to PIP2. They include: Kir, inward rectifier K+ channel; Kv, voltage-gated K+ channel; HERG, human ether-à-go-go-related gene K+ channel; HCN channel, hyperpolarization-activated, CNG, cyclic nucleotide-regulated channel; Cav, voltage-gated Ca2+ channel; TRP, transient receptor potential; TrpL, TRP-like; CNG, cyclic nucleotide-gated channel; ENaC, epithelial Na+ channel; NCX, Na+-Ca2+ exchanger; NHE, Na+-H+ exchanger; NME, Na+-Mg2+ exchanger; PMCA, plasma membrane Ca2+ ATPase. Activation means increase of open probability, prevention of run down, or recovery from desensitization. Shift g-V means that the conductance-voltage relation is shifted along the voltage axis by PIP2.
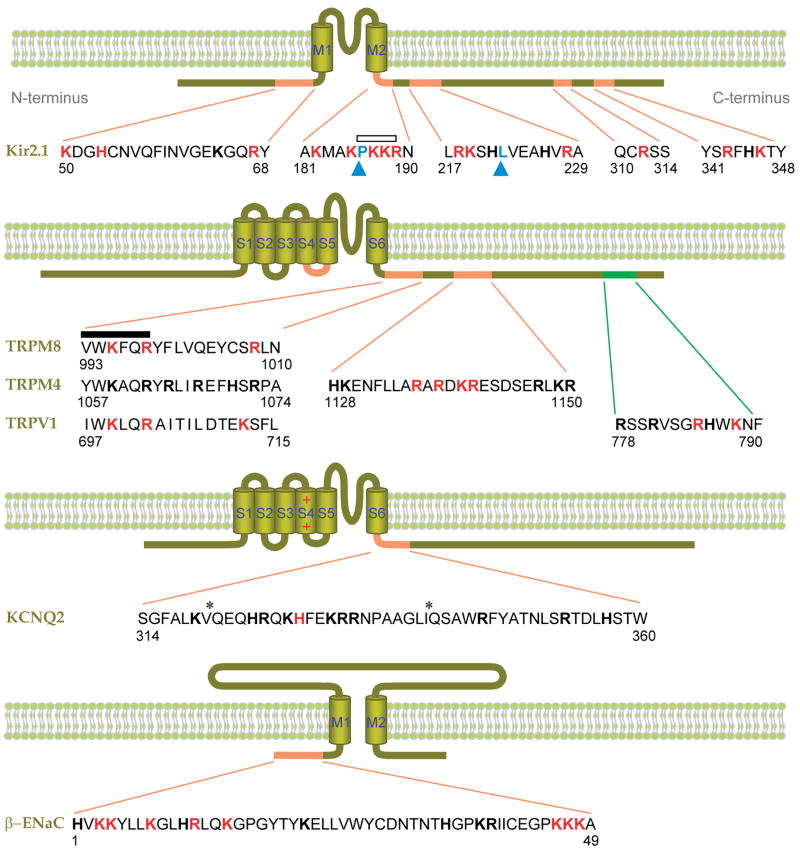
Putative PIP2-interacting residues of ion channels. Shown are Kir2.1 (32, 45), β-ENaC (48, 94), KCNQ2 (95), and several TRP channels (5, 54, 69, 64). Basic amino acids experimentally implicated in PIP2 binding are red, and other basic residues are shown in block bold. Hydrophobic residues implicated in PIP2 binding are indicated by blue arrowheads (76, 96). The distal C-terminal sequence of TRPV1 implicated in inhibition by PIP2 is green (64). Asterisks denote putative calmodulin binding sites. An open bar indicates a conserved PKKR domain in Kir channels involved in PIP2 binding, and the closed bar indicates the highly conserved TRP box (XWK(F/X)QR). Numbers indicate the amino acid positions. M1–2 and S1–6 denote the transmembrane domains.
References
-
- Baukrowitz T, Schulte U, Oliver D, Herlitze S, Krauter T, et al. PIP2 and PIP as determinants for ATP inhibition of KATP channels. Science. 1998;282:1141–4. - PubMed
-
- Bian J, Cui J, McDonald TV. HERG K+ channel activity is regulated by changes in phosphatidylinositol 4,5-bisphosphate. Circ Res. 2001;89:1168–76. - PubMed
-
- Bian JS, Kagan A, McDonald TV. Molecular analysis of PIP2 regulation of HERG and IKr. Am J Physiol Heart Circ Physiol. 2004;287:H2154–63. - PubMed
-
- Branstrom R, Leibiger IB, Leibiger B, Corkey BE, Berggren PO, et al. Long chain coenzyme A esters activate the pore-forming subunit (Kir6.2) of the ATP-regulated potassium channel. J Biol Chem. 1998;273:31395–400. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
