

A sensor histidine kinase co-ordinates cell wall architecture with cell division in Bacillus subtilis
- PMID: 18573169
- PMCID: PMC2574549
- DOI: 10.1111/j.1365-2958.2008.06308.x
A sensor histidine kinase co-ordinates cell wall architecture with cell division in Bacillus subtilis
Abstract
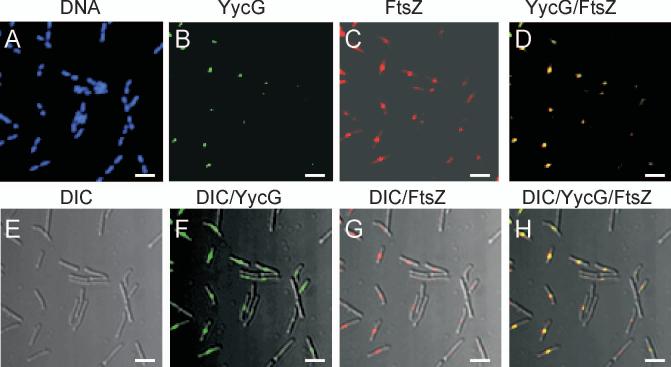
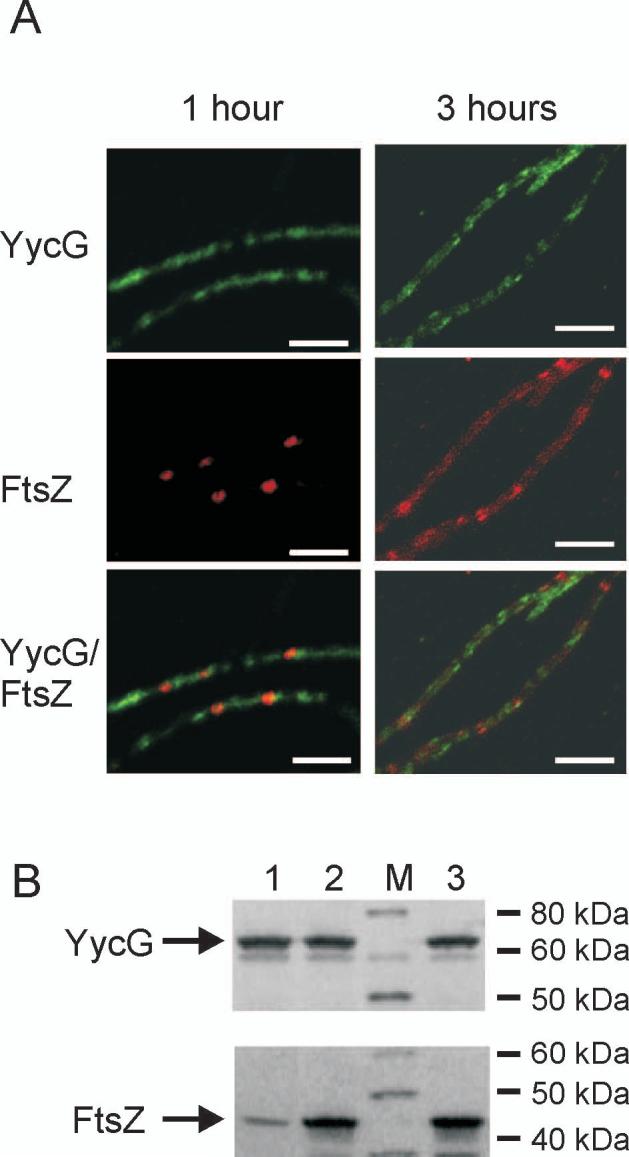
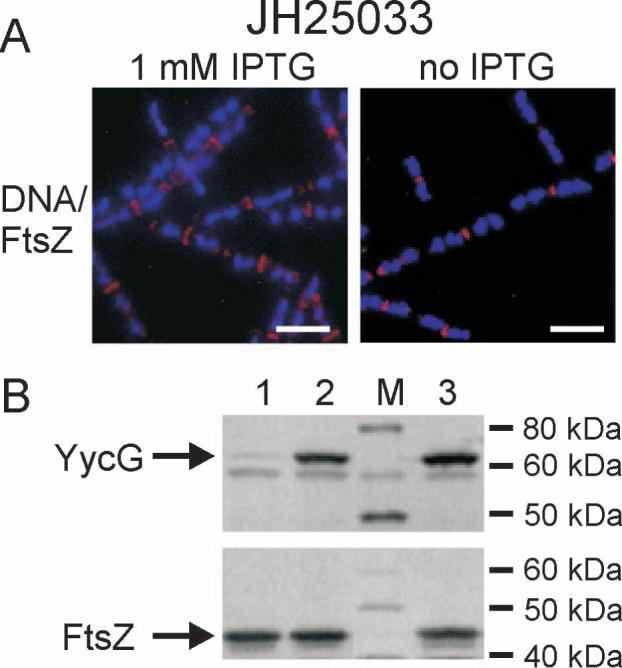
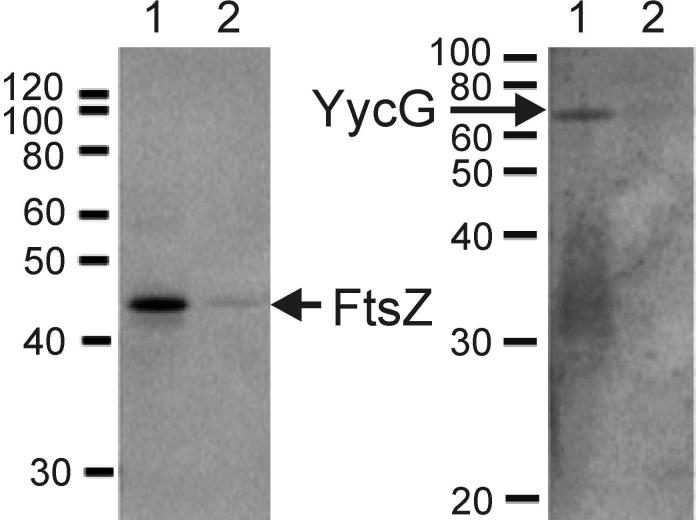
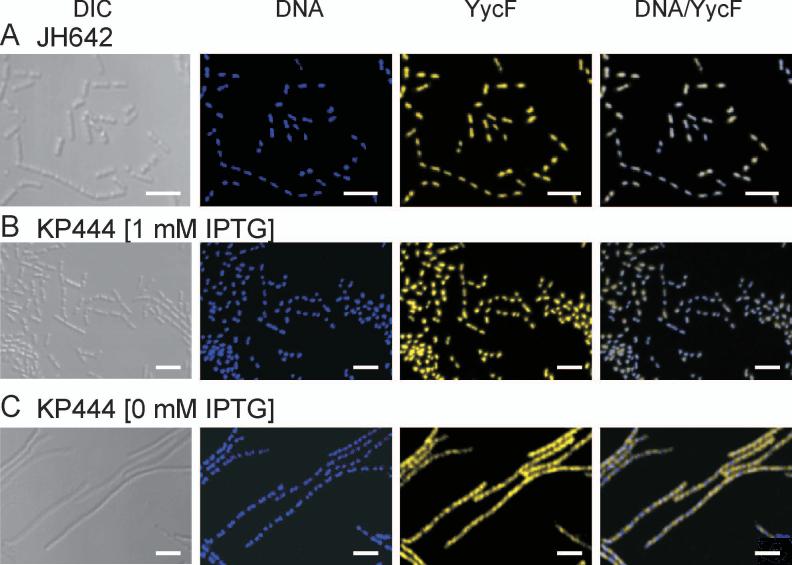
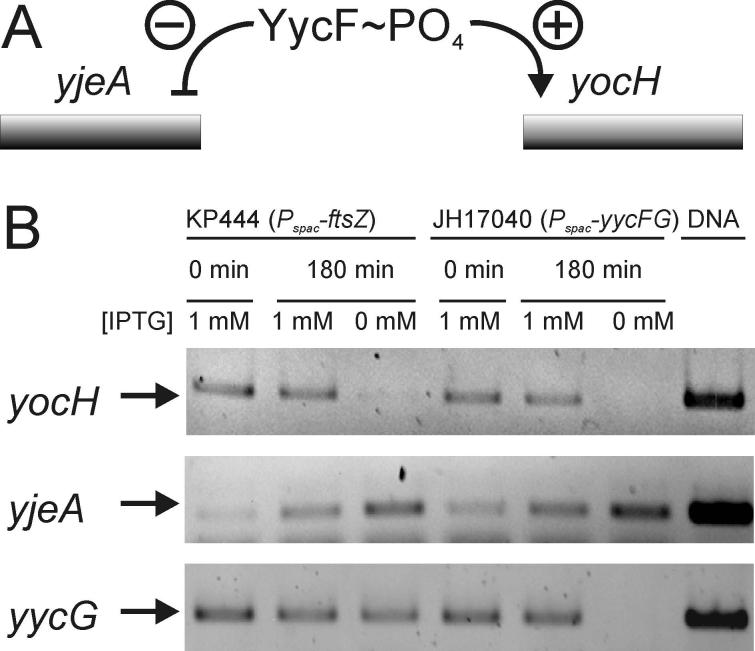
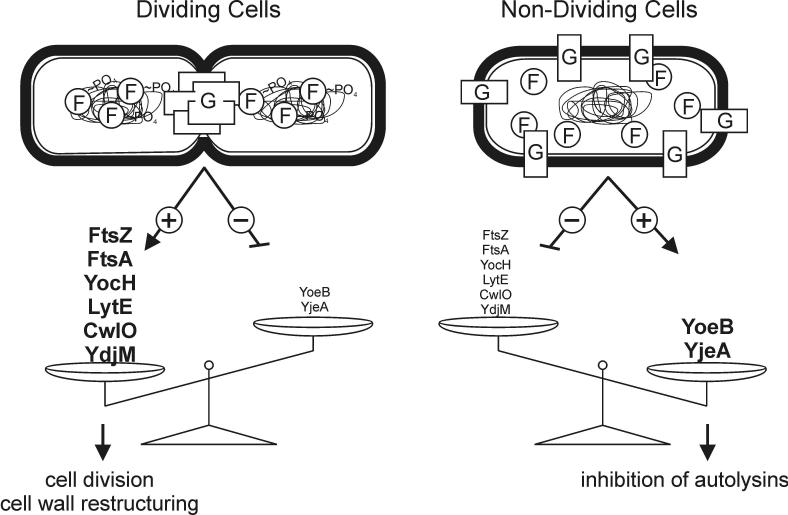
The concerted interconnection between processes driving DNA synthesis, division septum formation and cell wall synthesis and remodelling in rapidly growing bacteria requires precise co-ordination by signalling mechanisms that are, for the most part, unknown. The YycG (sensor histidine kinase)-YycF (response regulator/transcription factor) two-component system of Bacillus subtilis controls the synthesis of enzymes and their inhibitors that function in cell wall remodelling and cell separation. Here it is shown that the YycG sensor histidine kinase is a component of the division septum in growing cells. RT-PCR quantification of YycF approximately PO(4)-regulated gene transcription, in wild type and FtsZ-depleted, septum-less cells, indicated that YycG kinase activity on YycF is dependent on YycG localization to a division septum. The data support a model in which the YycG sensor kinase perceives information at the division septum and regulates the reciprocal synthesis of autolysins and autolysin inhibitors to co-ordinate growth and division with cell wall restructuring.
Figures
References
-
- Baker MD, Wolanin PM, Stock JB. Signal transduction in bacterial chemotaxis. Bioessays. 2006;28:9–22. - PubMed
-
- Beall B, Lutkenhaus J. FtsZ in Bacillus subtilis is required for vegetative septation and for asymmetric septation during sporulation. Genes Dev. 1991;5:447–455. - PubMed
-
- Bi EF, Lutkenhaus J. FtsZ ring structure associated with division in Escherichia coli. Nature. 1991;354:161–164. - PubMed
-
- Biondi EG, Reisinger SJ, Skerker JM, Arif M, Perchuk BS, Ryan KR, Laub MT. Regulation of the bacterial cell cycle by an integrated genetic circuit. Nature. 2006;444:899–904. - PubMed
-
- Bisicchia P, Noone D, Lioliou E, Howell A, Quigley S, Jensen T, Jarmer H, Devine KM. The essential YycFG two-component system controls cell wall metabolism in Bacillus subtilis. Mol Microbiol. 2007;65:180–200. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
