

IL-12- and IL-23-modulated T cells induce distinct types of EAE based on histology, CNS chemokine profile, and response to cytokine inhibition
- PMID: 18573909
- PMCID: PMC2442630
- DOI: 10.1084/jem.20080159
IL-12- and IL-23-modulated T cells induce distinct types of EAE based on histology, CNS chemokine profile, and response to cytokine inhibition
Abstract
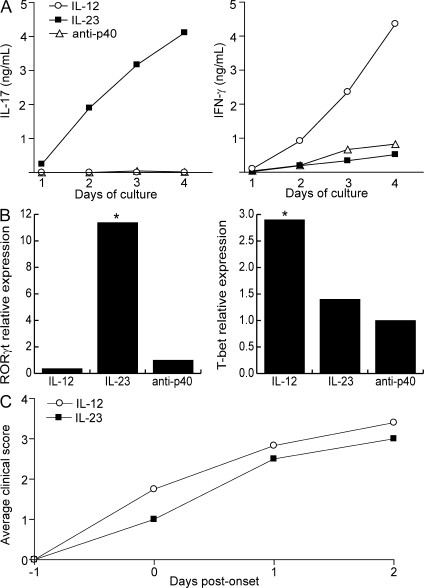
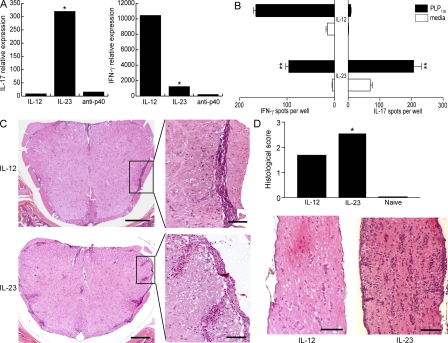
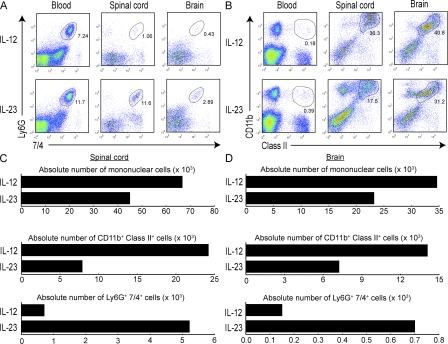
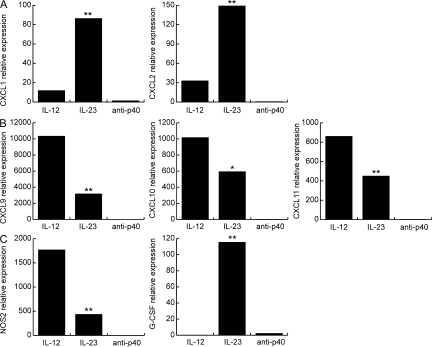
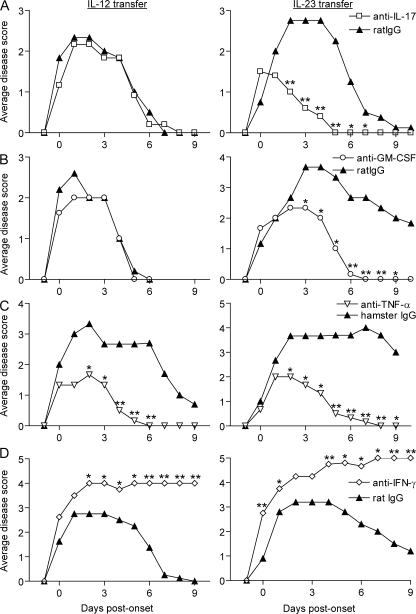
The interleukin (IL)-12p40 family of cytokines plays a critical role in the development of experimental autoimmune encephalomyelitis (EAE). However, the relative contributions of IL-12 and IL-23 to the pathogenic process remain to be elucidated. Here, we show that activation of uncommitted myelin-reactive T cells in the presence of either IL-12p70 or IL-23 confers encephalogenicity. Adoptive transfer of either IL-12p70- or IL-23-polarized T cells into naive syngeneic hosts resulted in an ascending paralysis that was clinically indistinguishable between the two groups. However, histological and reverse transcription-polymerase chain reaction analysis of central nervous system (CNS) tissues revealed distinct histopathological features and immune profiles. IL-12p70-driven disease was characterized by macrophage-rich infiltrates and prominent NOS2 up-regulation, whereas neutrophils and granulocyte-colony-stimulating factor (CSF) were prominent in IL-23-driven lesions. The monocyte-attracting chemokines CXCL9, 10, and 11 were preferentially expressed in the CNS of mice injected with IL-12p70-modulated T cells, whereas the neutrophil-attracting chemokines CXCL1 and CXCL2 were up-regulated in the CNS of mice given IL-23-modulated T cells. Treatment with anti-IL-17 or anti-granulocyte/macrophage-CSF inhibited EAE induced by transfer of IL-23-polarized, but not IL-12p70-polarized, cells. These findings indicate that autoimmunity can be mediated by distinct effector populations that use disparate immunological pathways to achieve a similar clinical outcome.
Figures
Comment in
-
A rush to judgment on Th17.J Exp Med. 2008 Jul 7;205(7):1517-22. doi: 10.1084/jem.20072066. J Exp Med. 2008. PMID: 18591407 Free PMC article.
References
-
- Fassbender, K., A. Ragoschke, S. Rossol, A. Schwartz, O. Mielke, A. Paulig, and M. Hennerici. 1998. Increased release of interleukin-12p40 in MS: association with intracerebral inflammation. Neurology. 51:753–758. - PubMed
-
- Moldovan, I.R., R.A. Rudick, A.C. Cotleur, S.E. Born, J.C. Lee, M.T. Karafa, and C.M. Pelfrey. 2003. Interferon gamma responses to myelin peptides in multiple sclerosis correlate with a new clinical measure of disease progression. J. Neuroimmunol. 141:132–140. - PubMed
-
- van Boxel-Dezaire, A.H., S.C. Hoff, B.W. van Oosten, C.L. Verweij, A.M. Drager, H.J. Ader, J.C. van Houwelingen, F. Barkhof, C.H. Polman, and L. Nagelkerken. 1999. Decreased interleukin-10 and increased interleukin-12p40 mRNA are associated with disease activity and characterize different disease stages in multiple sclerosis. Ann. Neurol. 45:695–703. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
