

Beta-caryophyllene is a dietary cannabinoid
- PMID: 18574142
- PMCID: PMC2449371
- DOI: 10.1073/pnas.0803601105
Beta-caryophyllene is a dietary cannabinoid
Abstract
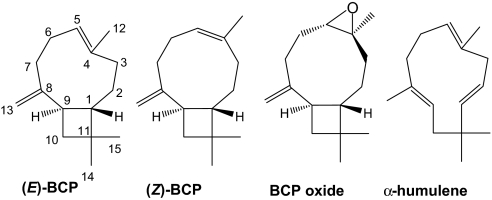
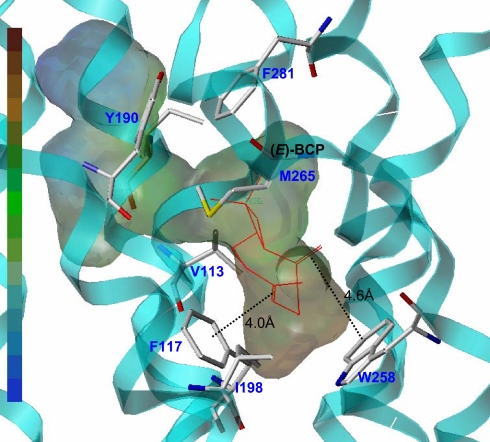
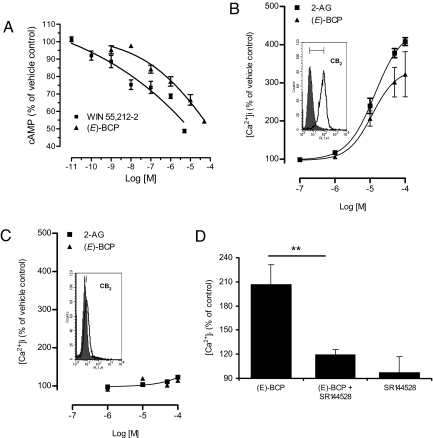
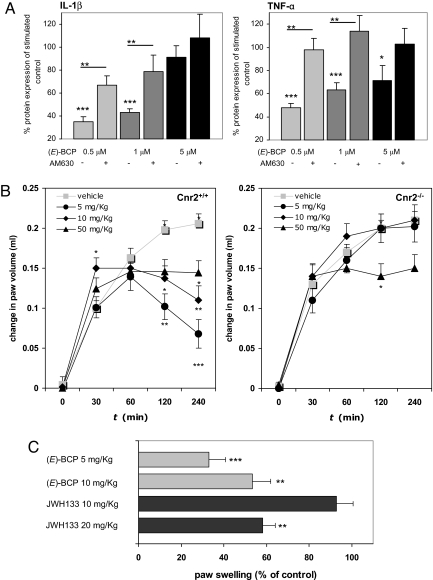
The psychoactive cannabinoids from Cannabis sativa L. and the arachidonic acid-derived endocannabinoids are nonselective natural ligands for cannabinoid receptor type 1 (CB(1)) and CB(2) receptors. Although the CB(1) receptor is responsible for the psychomodulatory effects, activation of the CB(2) receptor is a potential therapeutic strategy for the treatment of inflammation, pain, atherosclerosis, and osteoporosis. Here, we report that the widespread plant volatile (E)-beta-caryophyllene [(E)-BCP] selectively binds to the CB(2) receptor (K(i) = 155 +/- 4 nM) and that it is a functional CB(2) agonist. Intriguingly, (E)-BCP is a common constituent of the essential oils of numerous spice and food plants and a major component in Cannabis. Molecular docking simulations have identified a putative binding site of (E)-BCP in the CB(2) receptor, showing ligand pi-pi stacking interactions with residues F117 and W258. Upon binding to the CB(2) receptor, (E)-BCP inhibits adenylate cylcase, leads to intracellular calcium transients and weakly activates the mitogen-activated kinases Erk1/2 and p38 in primary human monocytes. (E)-BCP (500 nM) inhibits lipopolysaccharide (LPS)-induced proinflammatory cytokine expression in peripheral blood and attenuates LPS-stimulated Erk1/2 and JNK1/2 phosphorylation in monocytes. Furthermore, peroral (E)-BCP at 5 mg/kg strongly reduces the carrageenan-induced inflammatory response in wild-type mice but not in mice lacking CB(2) receptors, providing evidence that this natural product exerts cannabimimetic effects in vivo. These results identify (E)-BCP as a functional nonpsychoactive CB(2) receptor ligand in foodstuff and as a macrocyclic antiinflammatory cannabinoid in Cannabis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Rasmann S, et al. Recruitment of entomopathogenic nematodes by insect-damaged maize roots. Nature. 2005;434:732–737. - PubMed
-
- Orav A, Stulova I, Kailas T, Müürisepp M. Effect of storage on the essential oil composition of Piper nigrum L. fruits of different ripening states. J Agric Food Chem. 2004;52:2582–2586. - PubMed
-
- Jayaprakasha GK, Jagan Mohan Rao L, Sakariah KK. Volatile constituents from Cinnamomum zeylanicum fruit stalks and their antioxidant activities. J Agric Food Chem. 2003;51:4344–4348. - PubMed
-
- Mockute D, Bernotiene G, Judzentiene A. The essential oil of Origanum vulgare L. ssp. vulgare growing wild in vilnius district (Lithuania) Phytochemistry. 2001;57:65–69. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
