

p21 Ras/impedes mitogenic signal propagation regulates cytokine production and migration in CD4 T cells
- PMID: 18577512
- PMCID: PMC2516986
- DOI: 10.1074/jbc.M804084200
p21 Ras/impedes mitogenic signal propagation regulates cytokine production and migration in CD4 T cells
Abstract
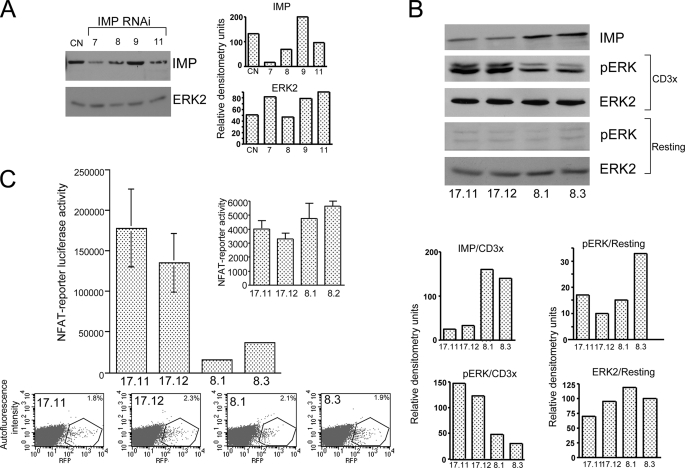
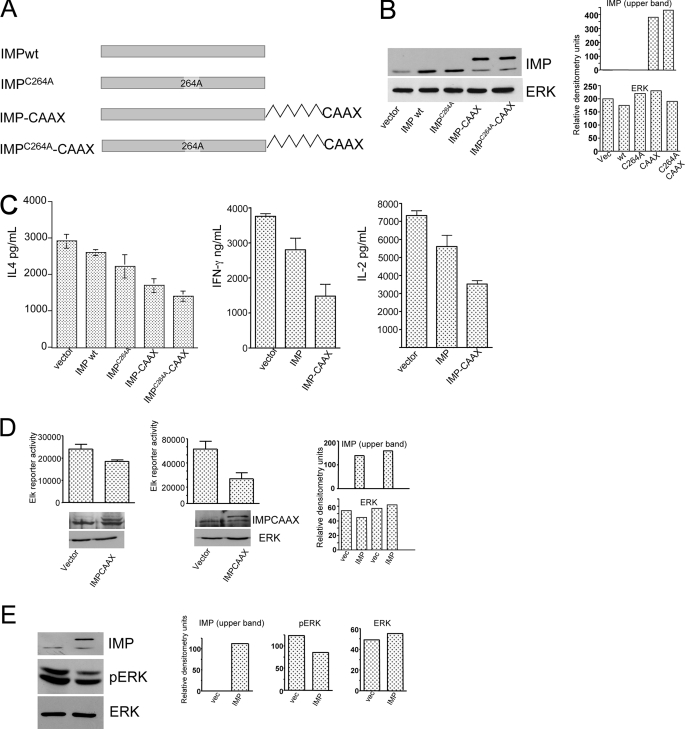
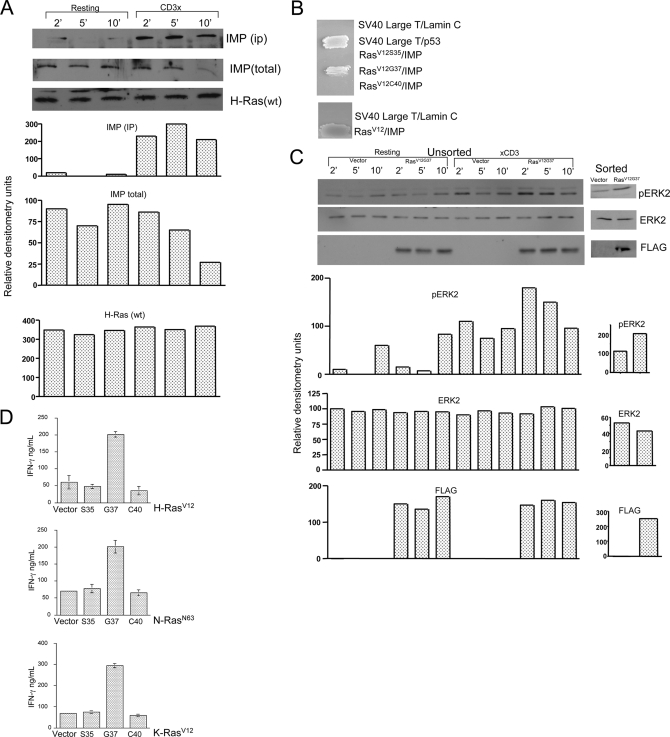
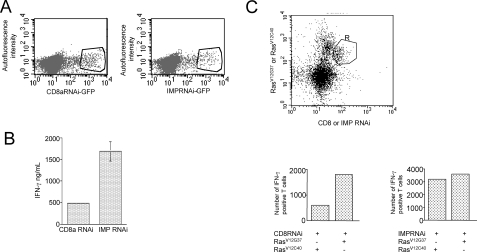
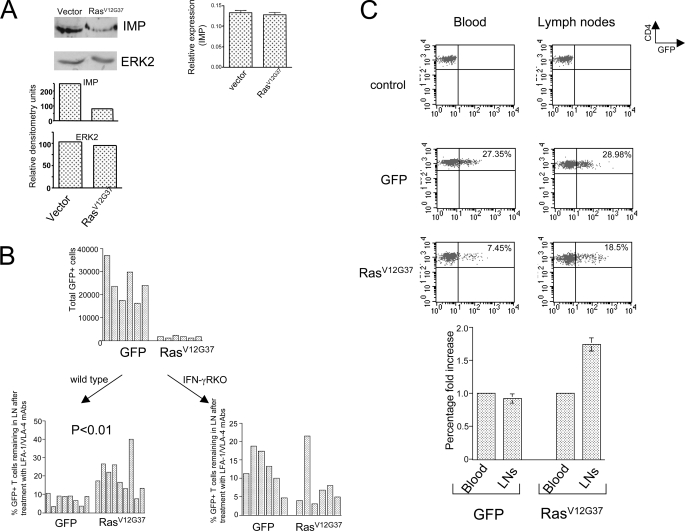
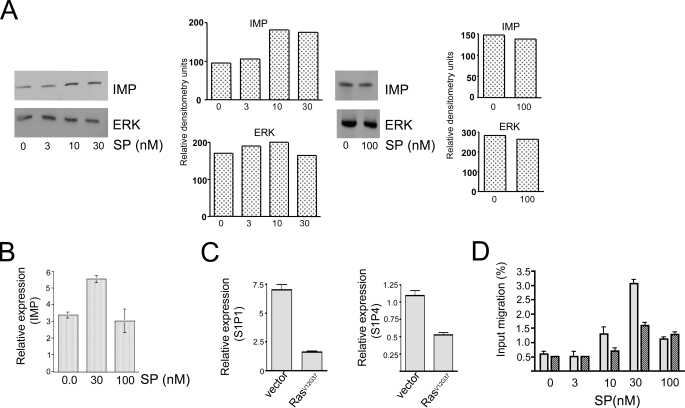
The propensity of T cells to generate coordinated cytokine responses is critical for the host to develop resistance to pathogens while maintaining the state of immunotolerance to self-antigens. The exact mechanisms responsible for preventing the overproduction of proinflammatory cytokines including interferon (IFN)-gamma are not fully understood, however. In this study, we examined the role of a recently described Ras GTPase effector and repressor of the Raf/MEK/ERK cascade called impedes mitogenic signal propagation (Imp) in limiting the induction of T-cell cytokines. We found that stimulation of the T cell receptor complex leads to the rapid development of a physical association between Ras and Imp. Consistent with the hypothesis that Imp inhibits signal transduction, we also found that disengagement of this molecule by the Ras(V12G37) effector loop mutant or RNA interference markedly enhances the activation of the NFAT transcription factor and IFN-gamma secretion. A strong output of IFN-gamma is responsible for the distinct lymphocyte traffic pattern observed in vivo because the transgenic or retroviral expression of Ras(V12G37) caused T cells to accumulate preferentially in the lymph nodes and delayed their escape from the lymphoid tissue, respectively. Together, our results describe a hitherto unrecognized negative regulatory role for Imp in the production of IFN-gamma in T cells and point to Ras-Imp binding as an attractive target for therapeutic interventions in conditions involving the production of this inflammatory cytokine.
Figures
Similar articles
-
Signaling threshold regulation by the Ras effector IMP.J Biol Chem. 2009 Apr 24;284(17):11007-11. doi: 10.1074/jbc.R800082200. Epub 2008 Dec 17. J Biol Chem. 2009. PMID: 19091743 Free PMC article. Review.
-
Ras regulates assembly of mitogenic signalling complexes through the effector protein IMP.Nature. 2004 Jan 15;427(6971):256-60. doi: 10.1038/nature02237. Nature. 2004. PMID: 14724641
-
Constitutive active p21ras enhances primary T cell responsiveness to Ca2+ signals without interfering with the induction of clonal anergy.Eur J Immunol. 2002 Sep;32(9):2500-9. doi: 10.1002/1521-4141(200209)32:9<2500::AID-IMMU2500>3.0.CO;2-S. Eur J Immunol. 2002. PMID: 12207334
-
Increased entry into the IFN-gamma effector pathway by CD4+ T cells selected by I-Ag7 on a nonobese diabetic versus C57BL/6 genetic background.J Immunol. 2001 Aug 1;167(3):1693-702. doi: 10.4049/jimmunol.167.3.1693. J Immunol. 2001. PMID: 11466393
-
Cytokine production by CD4+ and CD8+ T cells in mice following primary exposure to chemical allergens: evidence for functional differentiation of T lymphocytes in vivo.Int Arch Allergy Immunol. 1998 Jun;116(2):116-23. doi: 10.1159/000023934. Int Arch Allergy Immunol. 1998. PMID: 9652304
Cited by
-
Brap2 regulates temporal control of NF-κB localization mediated by inflammatory response.PLoS One. 2013;8(3):e58911. doi: 10.1371/journal.pone.0058911. Epub 2013 Mar 15. PLoS One. 2013. PMID: 23554956 Free PMC article.
-
E3 ubiquitin ligase GRAIL controls primary T cell activation and oral tolerance.Proc Natl Acad Sci U S A. 2009 Sep 29;106(39):16770-5. doi: 10.1073/pnas.0908957106. Epub 2009 Sep 17. Proc Natl Acad Sci U S A. 2009. PMID: 19805371 Free PMC article.
-
Signaling threshold regulation by the Ras effector IMP.J Biol Chem. 2009 Apr 24;284(17):11007-11. doi: 10.1074/jbc.R800082200. Epub 2008 Dec 17. J Biol Chem. 2009. PMID: 19091743 Free PMC article. Review.
-
Beyond the Cell Surface: Targeting Intracellular Negative Regulators to Enhance T cell Anti-Tumor Activity.Int J Mol Sci. 2019 Nov 20;20(23):5821. doi: 10.3390/ijms20235821. Int J Mol Sci. 2019. PMID: 31756921 Free PMC article. Review.
-
Central catalytic domain of BRAP (RNF52) recognizes the types of ubiquitin chains and utilizes oligo-ubiquitin for ubiquitylation.Biochem J. 2017 Sep 7;474(18):3207-3226. doi: 10.1042/BCJ20161104. Biochem J. 2017. PMID: 28768733 Free PMC article.
References
-
- Stokoe, D., MacDonald, S. G., Cadwallader, K., Symons, M., and Hancock, J. F. (1994) Science 264 1463–1467 - PubMed
-
- Vojtek, A. B., Hollenberg, S. M., and Cooper, J. A. (1993) Cell 74 205–214 - PubMed
-
- Downward, J., Graves, J. D., Warne, P. H., Rayter, S., and Cantrell, D. A. (1990) Nature 346 719–723 - PubMed
-
- Stefanova, I., Hemmer, B., Vergelli, M., Martin, R., Biddison, W. E., and Germain, R. N. (2003) Nat. Immunol. 4 248–254 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
