

Changes to gene expression associated with hybrid speciation in plants: further insights from transcriptomic studies in Senecio
- PMID: 18579474
- PMCID: PMC2607317
- DOI: 10.1098/rstb.2008.0080
Changes to gene expression associated with hybrid speciation in plants: further insights from transcriptomic studies in Senecio
Abstract
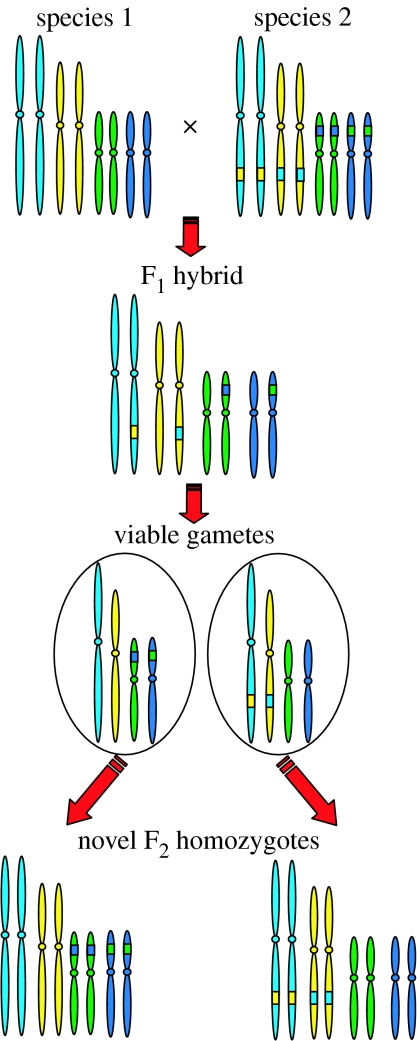
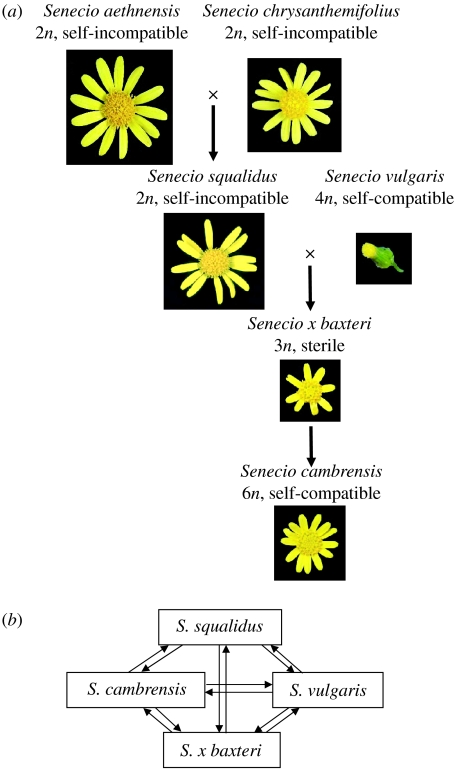
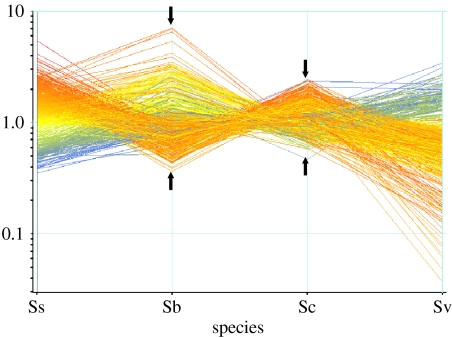
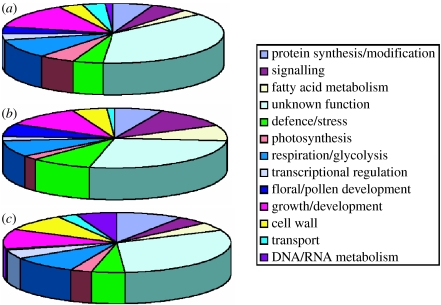
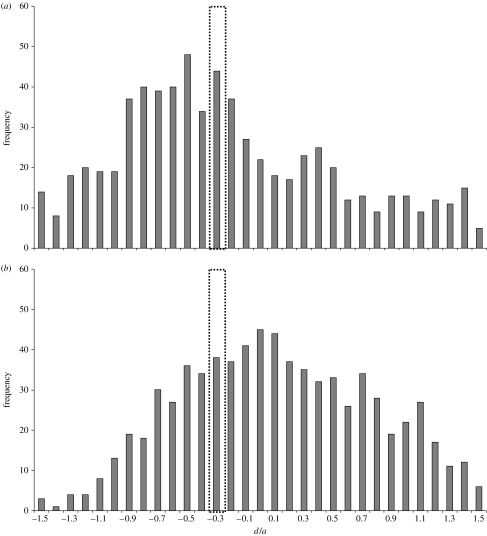
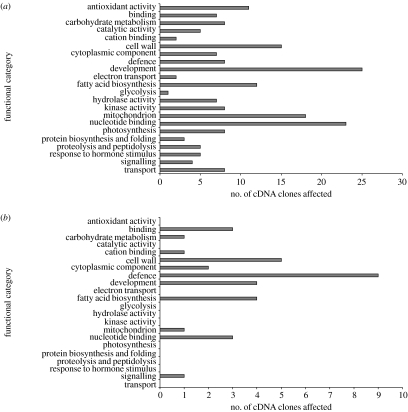
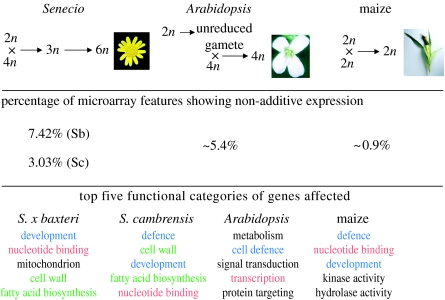
Interspecific hybridization is an important mechanism of speciation in higher plants. In flowering plants, hybrid speciation is usually associated with polyploidy (allopolyploidy), but hybrid speciation without genome duplication (homoploid hybrid speciation) is also possible, although it is more difficult to detect. The combination of divergent genomes within a hybrid can result in profound changes to both genome and transcriptome. Recent transcriptomic studies of wild and resynthesized homoploid and allopolyploid hybrids have revealed widespread changes to gene expression in hybrids relative to expression levels in their parents. Many of these changes to gene expression are 'non-additive', i.e. not simply the sum of the combined expression levels of parental genes. Some gene expression changes are far outside the range of gene expression in either parent, and can therefore be viewed as 'transgressive'. Such profound changes to gene expression may enable new hybrids to survive in novel habitats not accessible to their parent species. Here, we give a brief overview of hybrid speciation in plants, with an emphasis on genomic change, before focusing discussion on findings from recent transcriptomic studies. We then discuss our current work on gene expression change associated with hybrid speciation in the genus Senecio (ragworts and groundsels) focusing on the findings from a reanalysis of gene expression data obtained from recent microarray studies of wild and resynthesized allopolyploid Senecio cambrensis. These data, showing extensive non-additive and transgressive gene expression changes in Senecio hybrids, are discussed in the light of findings from other model systems, and in the context of the potential importance of gene expression change to hybrid speciation in plants.
Figures
References
-
- Abbott, R. J., Brennan, A. C., James, J. K., Forbes, D. G., Hegarty, M. J. & Hiscock, S. J. In press. Recent hybrid origin and invasion of the British Isles by a self-incompatible species, Oxford ragwort (Senecio squalidus L., Asteraceae). Biol. Inv
-
- Adams S, Vinkenoog R, Spielman M, Dickinson H.G, Scott R.J. Parent-of-origin effects on seed development in Arabidopsis thaliana require DNA methylation. Development. 2000;127:2493–2502. - PubMed
-
- Adams K.L, Cronn R, Percifield R, Wendel J.F. Genes duplicated by polyploidy show unequal contributions to the transcriptome and organ-specific reciprocal silencing. Proc. Natl Acad. Sci. USA. 2003;100:4649–4654. doi:10.1073/pnas.0630618100 - DOI - PMC - PubMed
-
- Adams K.L, Percifield R, Wendel J.F. Organ-specific silencing of genes in a newly synthesized cotton allotetraploid. Genetics. 2004;168:2217–2226. doi:10.1534/genetics.104.033522 - DOI - PMC - PubMed
-
- Alleman M, Doctor J. Genomic imprinting in plants: observations and evolutionary implications. Plant Mol. Biol. 2000;43:147–161. doi:10.1023/A:1006419025155 - DOI - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
