

Analgesic compound from sea anemone Heteractis crispa is the first polypeptide inhibitor of vanilloid receptor 1 (TRPV1)
- PMID: 18579526
- PMCID: PMC3259759
- DOI: 10.1074/jbc.M800776200
Analgesic compound from sea anemone Heteractis crispa is the first polypeptide inhibitor of vanilloid receptor 1 (TRPV1)
Abstract
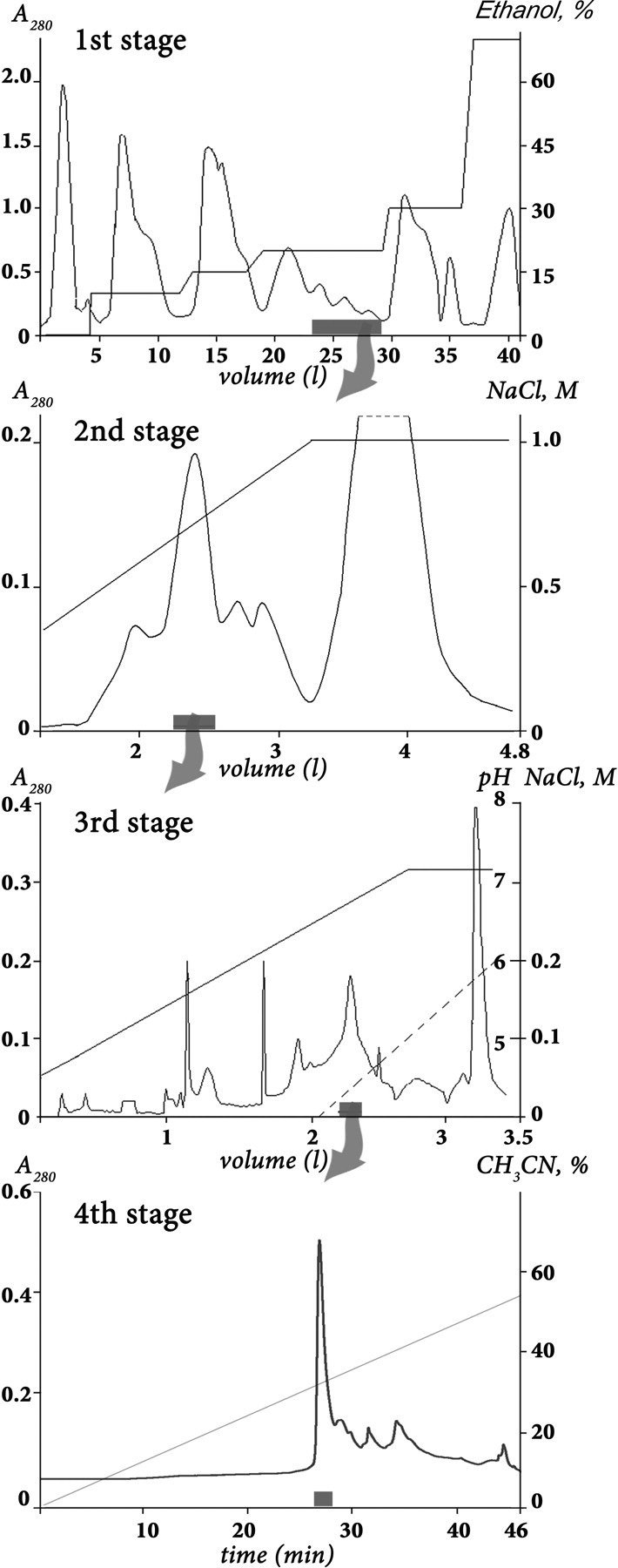
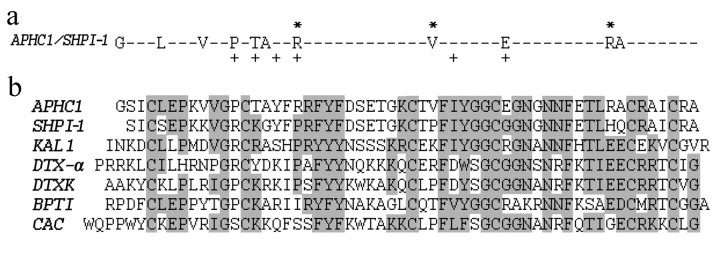
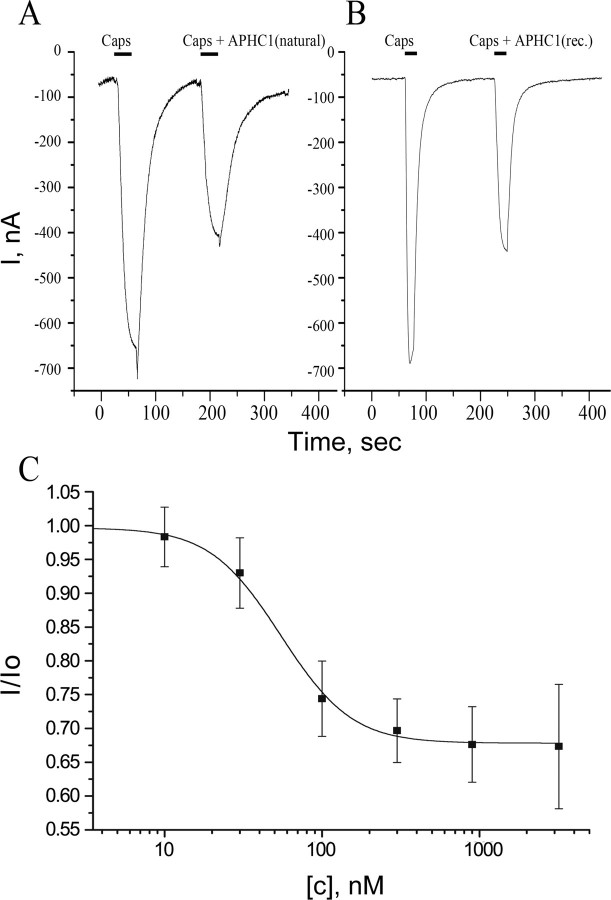
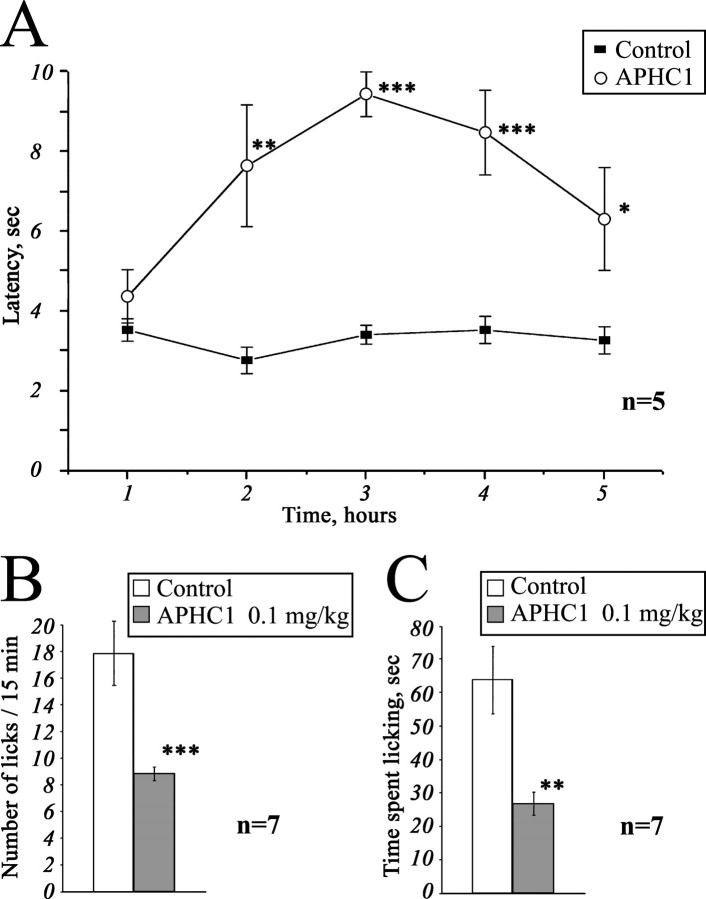
Venomous animals from distinct phyla such as spiders, scorpions, snakes, cone snails, or sea anemones produce small toxic proteins interacting with a variety of cell targets. Their bites often cause pain. One of the ways of pain generation is the activation of TRPV1 channels. Screening of 30 different venoms from spiders and sea anemones for modulation of TRPV1 activity revealed inhibitors in tropical sea anemone Heteractis crispa venom. Several separation steps resulted in isolation of an inhibiting compound. This is a 56-residue-long polypeptide named APHC1 that has a Bos taurus trypsin inhibitor (BPTI)/Kunitz-type fold, mostly represented by serine protease inhibitors and ion channel blockers. APHC1 acted as a partial antagonist of capsaicin-induced currents (32 +/- 9% inhibition) with half-maximal effective concentration (EC(50)) 54 +/- 4 nm. In vivo, a 0.1 mg/kg dose of APHC1 significantly prolonged tail-flick latency and reduced capsaicin-induced acute pain. Therefore, our results can make an important contribution to the research into molecular mechanisms of TRPV1 modulation and help to solve the problem of overactivity of this receptor during a number of pathological processes in the organism.
Figures

References
-
- Kawai, N., and Nakajima, T. (1993) in Natural and Synthetic Neurotoxins (Harvey, A. L., ed), pp. 319-345, Academic Press, London
-
- Miljanich, G. P. (1997) Sci. Med. (Phila.) 4 6-15
-
- Mortaria, M. R., Cunhaa, A. O., Ferreiraa, L. B., and Ferreira dos Santos, W. (2007) Pharmacol. Ther. 114 171-183 - PubMed
-
- Watters, M. R. (2005) Semin. Neurol. 25 278-289 - PubMed
-
- Guo, A., Vulchanova, L., Wang, J., Li, X., and Elde, R. (1999) Eur. J. Neurosci. 11 946-958 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
