

Renal Na+-K+-Cl- cotransporter activity and vasopressin-induced trafficking are lipid raft-dependent
- PMID: 18579701
- PMCID: PMC2536870
- DOI: 10.1152/ajprenal.90227.2008
Renal Na+-K+-Cl- cotransporter activity and vasopressin-induced trafficking are lipid raft-dependent
Abstract
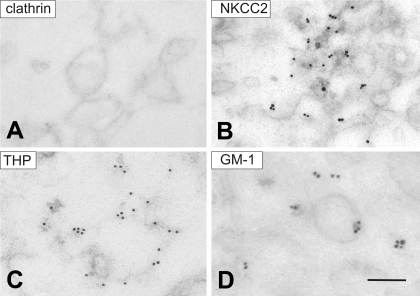
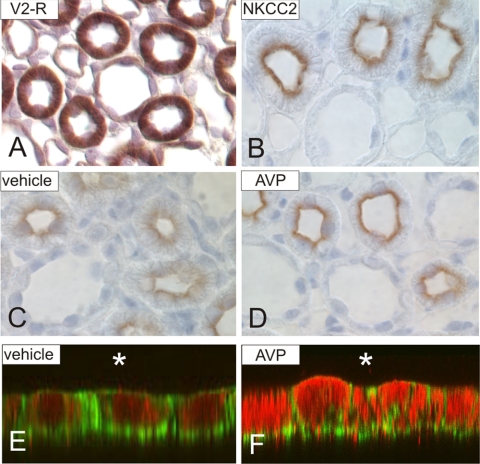
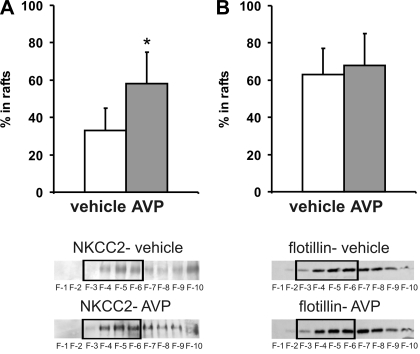
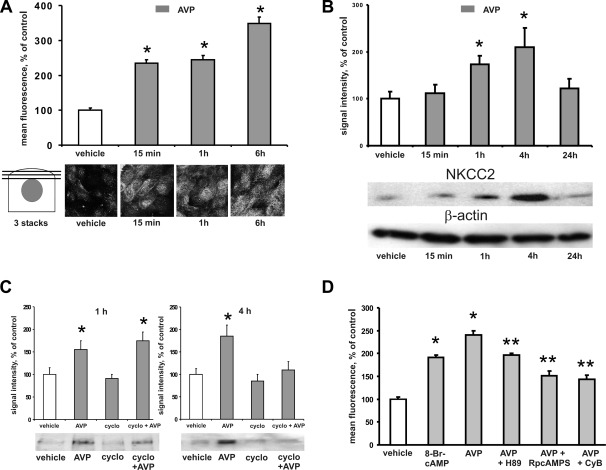
Apical bumetanide-sensitive Na(+)-K(+)-2Cl(-) cotransporter (NKCC2), the kidney-specific member of a cation-chloride cotransporter superfamily, is an integral membrane protein responsible for the transepithelial reabsorption of NaCl. The role of NKCC2 is essential for renal volume regulation. Vasopressin (AVP) controls NKCC2 surface expression in cells of the thick ascending limb of the loop of Henle (TAL). We found that 40-70% of Triton X-100-insoluble NKCC2 was present in cholesterol-enriched lipid rafts (LR) in rat kidney and cultured TAL cells. The related Na(+)-Cl(-) cotransporter (NCC) from rat kidney was distributed in LR as well. NKCC2-containing LR were detected both intracellularly and in the plasma membrane. Bumetanide-sensitive transport of NKCC2 as analyzed by (86)Rb(+) influx in Xenopus laevis oocytes was markedly reduced by methyl-beta-cyclodextrin (MbetaCD)-induced cholesterol depletion. In TAL, short-term AVP application induced apical vesicular trafficking along with a shift of NKCC2 from non-raft to LR fractions. In parallel, increased colocalization of NKCC2 with the LR ganglioside GM1 and their polar translocation were assessed by confocal analysis. Apical biotinylation showed twofold increases in NKCC2 surface expression. These effects were blunted by mevalonate-lovastatin/MbetaCD-induced cholesterol deprivation. Collectively, these findings demonstrate that a pool of NKCC2 distributes in rafts. Results are consistent with a model in which LR mediate polar insertion, activity, and AVP-induced trafficking of NKCC2 in the control of transepithelial NaCl transport.
Figures






Similar articles
-
Dynamin2, clathrin, and lipid rafts mediate endocytosis of the apical Na/K/2Cl cotransporter NKCC2 in thick ascending limbs.J Biol Chem. 2012 Nov 2;287(45):37824-34. doi: 10.1074/jbc.M112.386425. Epub 2012 Sep 12. J Biol Chem. 2012. PMID: 22977238 Free PMC article.
-
Molecular regulation of NKCC2 in the thick ascending limb.Am J Physiol Renal Physiol. 2011 Dec;301(6):F1143-59. doi: 10.1152/ajprenal.00396.2011. Epub 2011 Sep 7. Am J Physiol Renal Physiol. 2011. PMID: 21900458 Free PMC article. Review.
-
Constitutive endocytosis and recycling of NKCC2 in rat thick ascending limbs.Am J Physiol Renal Physiol. 2010 Nov;299(5):F1193-202. doi: 10.1152/ajprenal.00307.2010. Epub 2010 Aug 18. Am J Physiol Renal Physiol. 2010. PMID: 20719977 Free PMC article.
-
Activation of the bumetanide-sensitive Na+,K+,2Cl- cotransporter (NKCC2) is facilitated by Tamm-Horsfall protein in a chloride-sensitive manner.J Biol Chem. 2011 Aug 26;286(34):30200-10. doi: 10.1074/jbc.M111.222968. Epub 2011 Jul 7. J Biol Chem. 2011. PMID: 21737451 Free PMC article.
-
Trafficking and regulation of the NKCC2 cotransporter in the thick ascending limb.Curr Opin Nephrol Hypertens. 2017 Sep;26(5):392-397. doi: 10.1097/MNH.0000000000000351. Curr Opin Nephrol Hypertens. 2017. PMID: 28614115 Review.
Cited by
-
Dynamin2, clathrin, and lipid rafts mediate endocytosis of the apical Na/K/2Cl cotransporter NKCC2 in thick ascending limbs.J Biol Chem. 2012 Nov 2;287(45):37824-34. doi: 10.1074/jbc.M112.386425. Epub 2012 Sep 12. J Biol Chem. 2012. PMID: 22977238 Free PMC article.
-
Molecular regulation of NKCC2 in the thick ascending limb.Am J Physiol Renal Physiol. 2011 Dec;301(6):F1143-59. doi: 10.1152/ajprenal.00396.2011. Epub 2011 Sep 7. Am J Physiol Renal Physiol. 2011. PMID: 21900458 Free PMC article. Review.
-
SPAK differentially mediates vasopressin effects on sodium cotransporters.J Am Soc Nephrol. 2013 Feb;24(3):407-18. doi: 10.1681/ASN.2012040404. Epub 2013 Feb 7. J Am Soc Nephrol. 2013. PMID: 23393317 Free PMC article.
-
Adenylyl cyclase 6 enhances NKCC2 expression and mediates vasopressin-induced phosphorylation of NKCC2 and NCC.Am J Pathol. 2013 Jan;182(1):96-106. doi: 10.1016/j.ajpath.2012.09.014. Epub 2012 Nov 1. Am J Pathol. 2013. PMID: 23123217 Free PMC article.
-
Comprehensive Lipidome Profiling of the Kidney in Early-Stage Diabetic Nephropathy.Front Endocrinol (Lausanne). 2020 Jun 19;11:359. doi: 10.3389/fendo.2020.00359. eCollection 2020. Front Endocrinol (Lausanne). 2020. PMID: 32655493 Free PMC article.
References
-
- Amlal H, Legoff C, Vernimmen C, Paillard M, Bichara M. Na+-K+(NH4+)-2Cl− cotransport in medullary thick ascending limb: control by PKA, PKC, and 20-HETE. Am J Physiol Cell Physiol 271: C455–C463, 1996. - PubMed
-
- Bock J, Gulbins E. The transmembranous domain of CD40 determines CD40 partitioning into lipid rafts. FEBS Lett 534: 169–174, 2003. - PubMed
-
- Breton S, Lisanti MP, Tyszkowski R, McLaughlin M, Brown D. Basolateral distribution of caveolin-1 in the kidney. Absence from H+-ATPase-coated endocytic vesicles in intercalated cells. J Histochem Cytochem 46: 205–214, 1998. - PubMed
-
- Brown DA Lipid rafts, detergent-resistant membranes, and raft targeting signals. Physiology 21: 430–439, 2006. - PubMed
-
- Cavallone D, Malagolini N, Serafini-Cessi F. Mechanism of release of urinary Tamm-Horsfall glycoprotein from the kidney GPI-anchored counterpart. Biochem Biophys Res Commun 280: 110–114, 2001. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous