

MHC class I-specific inhibitory receptors and their ligands structure diverse human NK-cell repertoires toward a balance of missing self-response
- PMID: 18583565
- PMCID: PMC2532809
- DOI: 10.1182/blood-2008-03-143727
MHC class I-specific inhibitory receptors and their ligands structure diverse human NK-cell repertoires toward a balance of missing self-response
Abstract
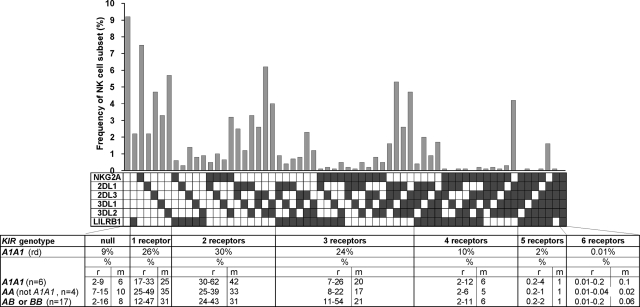
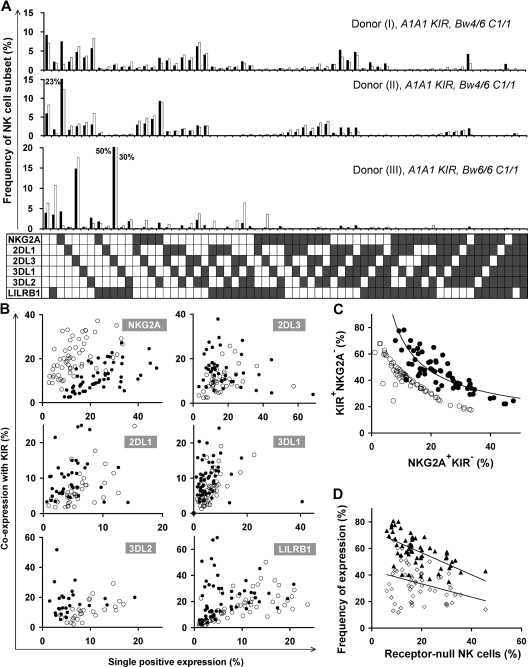
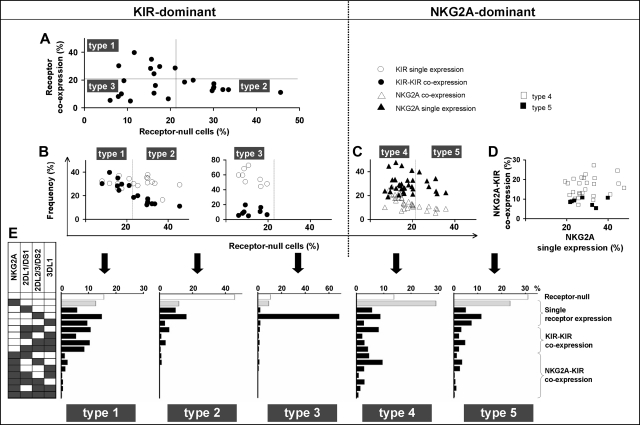
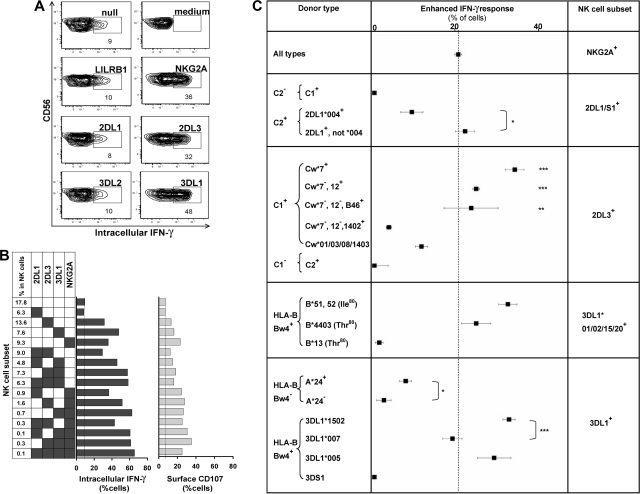
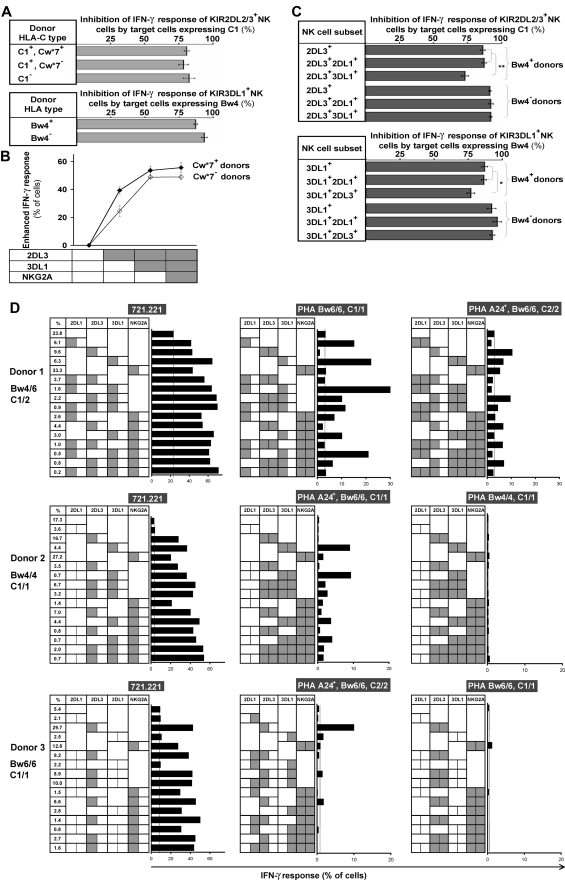
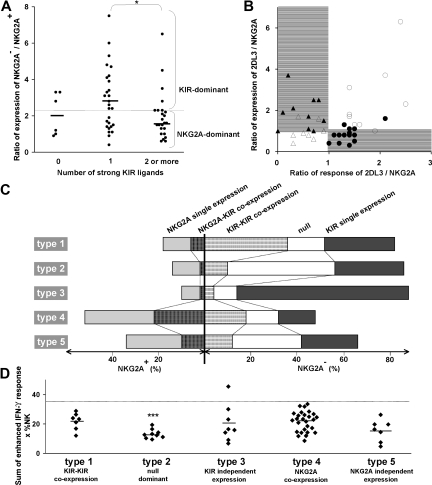
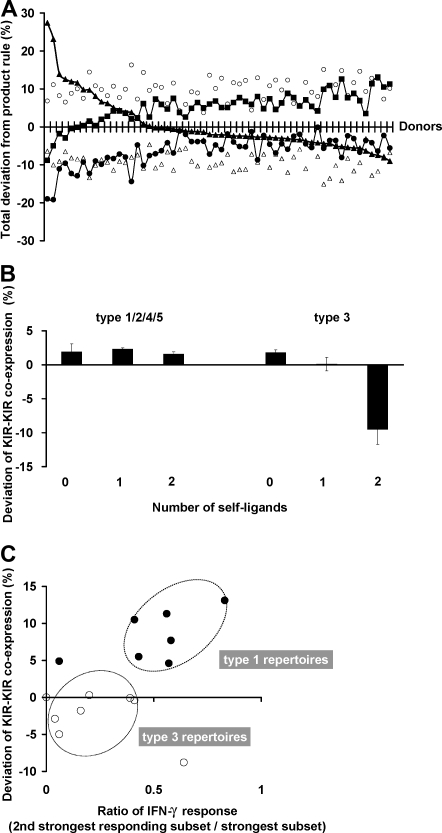
Variegated expression of 6 inhibitory HLA class I-specific receptors on primary NK cells was studied using high-dimension flow cytometry in 58 humans to understand the structure and function of NK-cell repertoires. Sixty-four subsets expressing all possible receptor com-binations were present in each repertoire, and the frequency of receptor-null cells varied among the donors. Enhancement in missing-self response between NK subsets varied substantially where subset responses were defined by donor KIR/HLA allotypes, reflecting the differences in interaction between inhibitory receptors and their ligands. This contrasted to the enhancement conferred by NKG2A, which was constant and of intermediate strength. We infer a mechanism that modulates frequencies of the NK subsets displaying diverse levels of missing-self response, a system that reduces the presence of KIR-expressing subsets that display either too strong or too weak a response and effectively replaces them with NKG2A-expressing cells in the repertoire. Through this high-resolution analysis of inhibitory receptor expression, 5 types of NK-cell repertoire were defined by their content of NKG2A(+)/NKG2A(-) cells, frequency of receptor-null cells, and degree of KIR receptor coexpression. The analyses provide new perspective on how personalized human NK-cell repertoires are structured.
Figures
References
-
- Parham P. MHC class I molecules and KIRs in human history, health and survival. Nat Rev Immunol. 2005;5:201–214. - PubMed
-
- Ruggeri L, Capanni M, Urbani E, et al. Effectiveness of donor natural killer cell alloreactivity in mismatched hematopoietic transplants. Science. 2002;295:2097–2100. - PubMed
-
- Karre K, Ljunggren HG, Piontek G, Kiessling R. Selective rejection of H-2-deficient lymphoma variants suggests alternative immune defence strategy. Nature. 1986;319:675–678. - PubMed
-
- Ljunggren HG, Karre K. In search of the “missing self”: MHC molecules and NK cell recognition. Immunol Today. 1990;11:237–244. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
